Le système urinaire (cours 2)
Le parenchyme rénal est subdivisé en deux parties,
le cortex et la médullaire; cette subdivision provenait
de la disposition des différents segments des néphrons.
Cette leçonci sera consacrée au glomérule
rénal.
 Les glomérules rénaux,
situés dans le cortex, sont dispersés entre les
tubules. Ce sont des corps globulaires d'un diamètre variant
entre 150 et 200 µm. Ils sont disposés en rangées
plus ou moins parallèles et perpendiculaires à la
capsule, le long des artères interlobulaires.
Les glomérules rénaux,
situés dans le cortex, sont dispersés entre les
tubules. Ce sont des corps globulaires d'un diamètre variant
entre 150 et 200 µm. Ils sont disposés en rangées
plus ou moins parallèles et perpendiculaires à la
capsule, le long des artères interlobulaires.
Le glomérule rénal est le premier segment du néphron.
Il est composé d'une touffe de capillaires en provenance
de l'artériole afférente et drainée par l'artériole
efférente. Cette touffe, le flocculus rénal,
est enveloppée par la capsule de Bowman. Entre les
capillaires du flocculus se trouve le mésangium.
Le glomérule
est une sphère dont la cavité est l'espace urinaire
de Bowman. Le flocculus y est suspendu. Les nombreuses anastomoses
entre ses capillaires le transforment en un vaste réseau.
Quelques cellules mésangiales ont été
dessinées en violet. Les deux feuillets qui délimitent
l'espace urinaire constituent la capsule de Bowman.
Le feuillet viscéral enveloppe très étroitement
la touffe vasculaire et en épouse tous les contours. Il
n'a été représenté que dans la partie
supérieure gauche du dessin et est en continuité
avec le feuillet externe ou pariétal. On appelle
pôle vasculaire l'endroit où l'artériole
afférente pénètre dans le glomérule
et où l'artériole efférente le quitte. A
ce niveau, le feuillet pariétal se réfléchit
dans le feuillet viscéral. Le pôle opposé
est appelé urinaire parce que la lumière
de la capsule de Bowman y est en continuité avec celle
du tube contourné proximal.
Dans une préparation histologique,
l'aspect des glomérules dépend de l'incidence de
la coupe. Le plus souvent la coupe est équatoriale et ne
passe donc par aucun des deux pôles. Dans ce cas, la lumière
de la capsule de Bowman a l'aspect d'un anneau continu
autour d'une masse centrale. Celleci se compose des capillaires
du flocculus entourés du feuillet viscéral. On peut,
dans la masse, distinguer les petites lumières des capillaires
et donc identifier certaines cellules endothéliales
désignées par une tête de flèche, parce
que leur noyau fait saillie dans ces lumières. Les noyaux
situés entre les capillaires appartiennent aux cellules
mésangiales. Le feuillet viscéral est difficilement
identifiable parce qu'il est étroitement accolé
aux capillaires. A certains endroits, on peut cependant repérer
le noyau plus clair d'une de ses cellules, parce qu'il fait saillie
dans l'espace urinaire (flèches). Les replis de ce feuillet
délimitent également les diverses incisures que
l'on observe dans la masse glomérulaire. Le feuillet pariétal
est un épithélium pavimenteux simple.
Parfois la coupe passe par le pôle vasculaire.
Dans ce cas, l'espace urinaire a toujours la forme d'un croissant
car le feuillet pariétal se réfléchit à
cet endroit et y est en continuité avec le feuillet viscéral.
Même quand la coupe passe par le pôle vasculaire,
l'aspect varie d'après son incidence. Ici, la coupe est
oblique et passe par une des artérioles, reconnaissable
à son endothélium et à sa tunique musculaire.
Lorsque la coupe intéresse le pôle vasculaire
mais que son incidence est à peu près perpendiculaire à la précédente,
on ne voit pas les artérioles afférente et efférente.
Par contre on distingue clairement un tube, dont la paroi est
composée de cellules cubiques, et un petit amas cellulaire
cunéiforme situé entre ce tube et le flocculus.
Le tube et l'amas cellulaire appartiennent à l'appareil
juxtaglomérulaire. La fente urinaire a la forme d'un croissant
parce que le feuillet viscéral de la capsule de Bowman
se réfléchit à cet endroit pour devenir feuillet
pariétal.
Le pôle urinaire
est à l'autre extrémité du glomérule.
Son identification est aisée car, à cet endroit,
la transition entre l'épithélium pavimenteux
du feuillet pariétal et l'épithélium cubique
du tube contourné proximal est brusque; de plus
la lumière de l'espace de Bowman est en continuité
avec celle du tube proximal. Les quelques agglomérats rouges
qui s'y trouvent sont de petits précipités protéiques
provoqués par la fixation.
Exceptionnellement l'incidence de la coupe à travers le
glomérule permet de voir les pôles urinaire (U) et vasculaire (V).
Dans ce dernier, on distingue les deux artérioles
(1 et 2) et l'appareil juxtaglomérulaire (3). L'espace
de Bowman est crescentiforme et en continuité avec la lumière
du tube proximal.
L'organisation des différents éléments
qui entrent dans la constitution du glomérule peut en coupe
être schématisée comme dans ce dessin. Le
glomérule possède un pôle vasculaire
(P.V.) et un pôle urinaire (P.U.). Les capillaires
du flocculus (C) sont représentés en vert.
Le feuillet viscéral (F.V.), représenté
en bleu, est une membrane continue qui cloisonne la masse des
capillaires et adhère étroitement à leur
paroi. Là où ils ne sont pas couverts par le feuillet
viscéral, les capillaires sont en rapport avec le mésangium,
composé de cellules noyées dans la substance mésangiale.
L'espace urinaire de Bowman (E.B.) est limité en
dehors par le feuillet pariétal (F.P.).
Voici cette organisation glomérulaire
telle qu'on peut l'observer sur une coupe d'un demi-micron colorée
au bleu de toluidine. On reconnaît aisément les capillaires
délimités par les cellules endothéliales
dont le noyau fait saillie dans la lumière, qui contient
parfois quelques érythrocytes. Le flocculus est bordé
en périphérie par les cellules du feuillet viscéral,
les podocytes dont nous reparlerons. Entre les anses capillaires
se trouve le mésangium dont les cellules sont noyées
dans la substance mésangiale amorphe (têtes de flèche).
L'endothélium des capillaires
est du type fenestré. Les expansions très
fines des cellules endothéliales sont percées de
nombreux pores dont le diamètre est compris entre 25 et
100 nm. Ils sont parfois munis d'un diaphragme dont
l'épaisseur ne dépasse pas 7 nm. Près du
noyau, la coupe de la paroi endothéliale est tangentielle,
les pores sont vus de face, ils forment un réseau complexe.
Les cellules mésangiales,
de forme irrégulière, présentent de nombreux
prolongements et contiennent les organites cytoplasmiques habituels.
Leur origine est très controversée. Pour certains,
elles seraient d'origine conjonctive; d'autres les assimilent
à des cellules musculaires; d'autres enfin leur
attribuent une origine monocytaire. Tous cependant admettent
qu'elles ont une grande activité de phagocytose,
fonction démontrée par l'injection de particules
marquées que l'on retrouve après un certain temps
dans ces cellules. On suppose qu'elles phagocytent les résidus
de filtration déposés sur la membrane basale. C'est
pourquoi certains ont émis l'hypothèse qu'elles
interviennent dans la régénération constante
de la membrane basale; la cellule mésangiale en éliminerait
les couches profondes les plus anciennes tandis que, dans le même
temps, les cellules du feuillet viscéral déposeraient
de nouvelles couches. Notons cependant que les cellules mésangiales
ont été longtemps considérées comme
les cellules de support des anses capillaires assurant la cohésion
du flocculus.
Les cellules mésangiales sont entourées d'une matrice
amorphe de densité variable, la substance mésangiale.
Sa quantité est considérablement augmentée
dans certains cas pathologiques, notamment lorsque s'y accumulent
des immunoglobulines. Leurs dépôts se traduisent
par des masses amorphes plus denses aux électrons que la
substance ellemême.
Le feuillet pariétal
ou feuillet capsulaire est particulièrement simple. Il
est constitué d'une couche unique de cellules pavimenteuses
qui, près du pôle vasculaire, se transforme en feuillet
viscéral. Ses cellules aplaties dont le noyau fait saillie
dans l'espace urinaire, deviennent plus hautes à mesure
que l'on se rapproche du pôle urinaire, finalement elles
acquièrent un aspect cubique et l'ultrastructure des cellules
épithéliales tubulaires.
Le feuillet viscéral
quant à lui est profondément modifié par
ses rapports immédiats avec les capillaires du flocculus.
Les cellules de ce feuillet sont des podocytes. La microscopie
électronique à balayage permet de mettre en évidence
la structure tridimensionnelle du feuillet viscéral. On
se trouve ici dans l'espace de Bowman. La masse centrale est un
corps cellulaire contenant le noyau. Elle émet des prolongements
relativement épais qui s'étalent de façon
radiaire en formant une corbeille autour du vaisseau. De ces prolongements
se détachent des diverticules plus courts. Ce sont les
pédicelles qui s'organisent avec ceux des podocytes
voisins tout en ménageant un espace, la fente de filtration.
Bien qu'épithéliales à l'origine, ces cellules
ont perdu, au cours de l'embryogenèse, certaines de leurs
caractéristiques épithéliales et notamment
celles qui leur permettaient de rester étroitement associées.
Un plus fort grossissement
de la partie encadrée dans la micrographie précédente
permet de préciser les rapports des pédicelles.
Ceuxci proviennent de prolongements primaires voisins qui
peuvent appartenir à plusieurs podocytes. Ils sont situés
dans des plans différents et s'enchevêtrent les uns
dans les autres, formant ainsi une sorte de pelote à la
surface du vaisseau. La fente de filtration est l'espace très
étroit et continu qui sépare les pédicelles
enchevêtrés.
La microscopie électronique à transmission
permet, à relativement faible grossissement, de préciser
les rapports entre les podocytes et les autres éléments
du glomérule. On reconnaît très facilement
le capillaire (C) délimité par un endothélium
fenestré. Appliqués contre le capillaire se trouvent
le corps d'un podocyte (P) et de très nombreux prolongements
qui s'engrènent les uns dans les autres. A certains endroits,
et surtout en périphérie, ces prolongements recouvrent
le mésangium représenté ici par une
cellule (M) entourée de substance mésangiale.
Les prolongements des podocytes
contiennent très peu d'organites mais un grand nombre de
microfilaments. Ce plus fort grossissement met mieux en
évidence la relation entre ces prolongements et les capillaires
dont ils ne sont séparés que par une membrane basale.
Cette membrane basale
est composée d'une couche centrale dense aux électrons
et de deux couches claires. Elle est apparemment unique mais,
puisque tout épithélium repose sur sa membrane basale
propre, elle résulte en fait de la fusion de deux membranes
distinctes, l'une produite par les cellules endothéliales
et l'autre par les podocytes. Cette double origine
est confirmée par l'analyse histochimique des glycosaminoglycanes
qui la composent et par le fait que, dans certaines conditions
pathologiques, elle se scinde en deux feuillets distincts.
Il existe donc entre la lumière du vaisseau et la lumière
de la capsule une barrière composée de trois couches:
l'endothélium vasculaire, la membrane basale et l'épithélium
de la capsule de Bowman. Une seule, la membrane basale, est
continue. D'un point de vue physiologique cette barrière
est la membrane de filtration.
Cette membrane est semiperméable; elle permet le
passage de l'eau et des petites molécules mais empêche
celui des molécules de poids moléculaire supérieur
à 68.000 correspondant à celui de l'albumine. L'endothélium
vasculaire, vu la taille de ses pores cytoplasmiques, ne peut
constituer un filtre efficace; on attribue par conséquent
la semiperméabilité de la membrane de filtration
à la membrane basale.
La filtration glomérulaire est la première étape
de la formation de l'urine. Elle est complexe mais peut de prime
abord être assimilée à une ultrafiltration
du plasma à travers la membrane de filtration. L'urine
primitive, que l'on peut analyser en la prélevant dans
la lumière de la capsule de Bowman au moyen d'une micropipette,
a le même pH, la même pression osmotique, la même
concentration en sels et en éléments organiques
que le plasma déprotéiné. Elle ne contient
aucun des constituants cellulaires du sang et aucune protéine
sanguine de poids moléculaire élevé.
Chaque minute l'ultrafiltration
produit 125 ml d'urine primitive qui représentent environ
le cinquième du volume plasmatique passant dans
les reins. Son importance dépend théoriquement des
pressions hydrostatiques, osmotiques et oncotiques qui règnent
dans les compartiments vasculaire et urinaire. La pression hydrostatique
utile est de 75 mm Hg, soit la différence entre la pression
hydrostatique des capillaires (90 mm Hg) et la pression hydrostatique
de la capsule de Bowman (15 mm Hg). La pression osmotique
est identique dans les deux compartiments puisqu'elle est due
aux sels et à d'autres substances de petite taille moléculaire
qui se trouvent à concentration égale dans le plasma
et dans l'urine primitive. La pression oncotique, due aux
protéines, est de 30 mm Hg dans le plasma; elle est nulle
dans l'urine primitive. La pression qui permet l'ultrafiltration
à travers la membrane de filtration est donc égale
à 45 mm Hg.
Nous avons terminé ainsi l'étude du glomérule
rénal, première partie du néphron dont la
fonction unique est la filtration du plasma. Le cours suivant
sera consacré aux segments tubulaires.
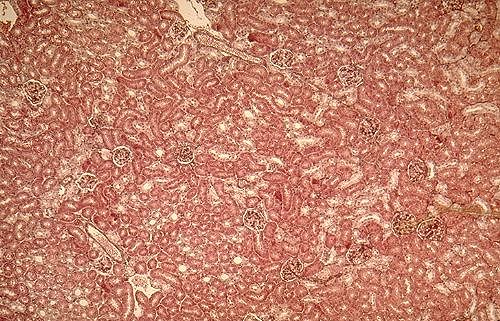 Les glomérules rénaux,
situés dans le cortex, sont dispersés entre les
tubules. Ce sont des corps globulaires d'un diamètre variant
entre 150 et 200 µm. Ils sont disposés en rangées
plus ou moins parallèles et perpendiculaires à la
capsule, le long des artères interlobulaires.
Les glomérules rénaux,
situés dans le cortex, sont dispersés entre les
tubules. Ce sont des corps globulaires d'un diamètre variant
entre 150 et 200 µm. Ils sont disposés en rangées
plus ou moins parallèles et perpendiculaires à la
capsule, le long des artères interlobulaires.