Le système urinaire (cours 1)
Le système urinaire regroupe l'ensemble des organes nécessaires
à l'élaboration, au transport, au stockage et à
l'élimination de l'urine. Il intervient avec d'autres organes
dans l'équilibre hydrique et ionique du milieu intérieur.
Il comprend les reins (1) qui élaborent l'urine,
et les voies urinaires, ensemble de canaux excréteurs
et de réservoirs, parmi lesquels on distingue les calices
(2), les bassinets (3), les uretères (4),
la vessie (5) et l'urètre (6).
 De teinte rougebrun et de consistance ferme, les reins
ont la forme d'un haricot. La face antérieure, visible
sur cette diapositive, est bombée et la face postérieure
est plane. Le bord externe est convexe, et l'interne est échancré.
A ce niveau se trouve le hile, comblé d'un tissu
graisseux de teinte jaunâtre et par où passent les
vaisseaux, les nerfs et l'uretère. L'extrémité
supérieure est plus large que l'inférieure. Chez
l'adulte, les reins mesurent environ 12 cm de haut, 6 cm de large
et 3 cm en épaisseur. Le poids individuel des reins peut
varier, mais le poids global est généralement voisin
de 300 g.
De teinte rougebrun et de consistance ferme, les reins
ont la forme d'un haricot. La face antérieure, visible
sur cette diapositive, est bombée et la face postérieure
est plane. Le bord externe est convexe, et l'interne est échancré.
A ce niveau se trouve le hile, comblé d'un tissu
graisseux de teinte jaunâtre et par où passent les
vaisseaux, les nerfs et l'uretère. L'extrémité
supérieure est plus large que l'inférieure. Chez
l'adulte, les reins mesurent environ 12 cm de haut, 6 cm de large
et 3 cm en épaisseur. Le poids individuel des reins peut
varier, mais le poids global est généralement voisin
de 300 g.
Les deux reins
sont rétropéritonéaux et appliqués
contre la paroi postérieure de l'abdomen, de part et d'autre
de la colonne vertébrale. Le hile du rein droit (1) est
situé au niveau de la deuxième vertèbre lombaire.
Du fait de ses rapports avec le foie non représenté
sur ce schéma, il est situé 2 cm plus bas que le
rein gauche (2). Le rein droit est en rapport avec le duodénum,
la tête du pancréas et l'angle colique droit. Le
rein gauche est en rapport avec la rate, la queue du pancréas
et l'angle colique gauche.
Chaque rein est entouré de 2 capsules:
une capsule fibreuse qui lui est propre (1) et une capsule
fibroadipeuse externe. La première adhère
intimement au parenchyme rénal. La seconde délimite
la loge rénale et maintient les reins en place. Elle est
constituée d'une lame fibreuse externe (2) et d'une couche
graisseuse (3) qui renferme des vaisseaux. Les deux capsules sont
représentées ici en coupe frontale et du côté
droit. La loge rénale est fixée au diaphragme (4)
par un feutrage fibreux très dense. A son pôle caudal,
la loge est ouverte et ceci explique la possibilité d'une
descente du rein, encore appelée ptose rénale. Audessus
du rein, se trouve la surrénale qui en est séparée
par un dédoublement du fascia fibreux.
En examinant le schéma
d'une section sagittale médiane, on remarque que le hile
se prolonge à l'intérieur du rein par une cavité
appelée le sinus rénal. Cette cavité
qui contient le début des voies urinaires, les gros vaisseaux
et les nerfs, est comblée par des expansions de la capsule
fibroadipeuse. Le parenchyme est composé de deux
grandes parties: une zone externe claire, la corticale
(C), de couleur brunrouge et une zone interne plus sombre,
la médullaire (M). La médullaire est constituée
de structures coniques, les pyramides de Malpighi (1) entre
lesquelles s'insinuent les colonnes de Bertin (2), prolongements
du cortex rénal contenant des vaisseaux en provenance du
sinus. Dans l'espèce humaine, le nombre des pyramides varie
entre 8 et 18; dans d'autres espèces, comme le rat par
exemple, il n'en existe qu'une. Le sommet de chaque pyramide est
appelé papille (3) et sa pointe est percée
de petits orifices, les pores urinaires, points d'émergence
des tubes rénaux. C'est par là que l'urine quitte
le parenchyme rénal pour s'écouler dans les voies
urinaires. Chaque papille est enveloppée par l'épithélium
du calice (4), début des voies urinaires dont nous
détaillerons l'agencement ultérieurement. La transition
entre les pyramides et le cortex est marquée par les stries
de Ferrein (5).
Voici une tranche sagittale
comparable à celle du schéma précédent.
La distinction entre cortex et médullaire est évidente.
La médullaire est partagée en pyramides. Le sommet
de chaque pyramide, ou papille rénale, fait saillie dans
le sinus rénal, comblé par du tissu adipeux. Les
pyramides sont séparées les unes des autres par
des expansions du cortex et du sinus, les colonnes de Bertin.
 Le parenchyme rénal peut donc être subdivisé
en deux grandes parties, le cortex et la médullaire.
Elles sont, d'un point de vue histologique, très différentes
et, avant de décrire le détail de chaque structure
rénale, nous examinerons leur aspect général
à faible grossissement.
Le parenchyme rénal peut donc être subdivisé
en deux grandes parties, le cortex et la médullaire.
Elles sont, d'un point de vue histologique, très différentes
et, avant de décrire le détail de chaque structure
rénale, nous examinerons leur aspect général
à faible grossissement.
Dans le parenchyme cortical se trouvent de nombreux petits amas
ronds dispersés; ce sont les glomérules de Malpighi.
Ceuxci, typiques du cortex, sont néanmoins absents
de sa zone la plus externe (1) située immédiatement
sous la capsule fibreuse. Dans le reste du cortex (2) et
en dehors des glomérules, le parenchyme est constitué
d'un lacis de tubes dont les plus nombreux sont très colorés.
La zone (3), située en dessous de la ligne pointillée,
est la médullaire.
Le parenchyme de la médullaire,
subdivisé en pyramides, est composé exclusivement
de tubes qui, dans cette préparation, sont moins
colorés que les tubes corticaux. Leur diamètre est
très variable, certains sont larges et d'autres sont étroits.
La base des pyramides
se prolonge par des expansions perpendiculaires à la
surface rénale qui correspondent aux striations intermédiaires
entre le cortex et la médullaire décrites dans la
coupe macroscopique. Elles sont appelées pyramides de
Ferrein ou stries médullaires. Cette région
est une zone de transition où les tubes passent du cortex
dans la médullaire ou inversement.
Au niveau des papilles
qui font saillie dans le sinus rénal, le parenchyme de
la médullaire est composé de tubes plus volumineux
dont le diamètre est à peu près constant.
En fonction de sa microanatomie, le rein a été subdivisé
en lobes et en lobules. Un lobe est l'équivalent
d'une pyramide et du territoire cortical adjacent.
Un lobule est l'équivalent d'une strie médullaire
et du territoire cortical qui l'entoure.
Les glomérules de Malpighi, situés dans le cortex,
et tous les tubes tant corticaux que médullaires sont des
segments des unités histophysiologiques du rein, les néphrons.
Il en existe un million à un million quatre cent mille
par rein. Bien que leur structure générale soit
identique, ils diffèrent selon leur taille, leur localisation
et certaines particularités fonctionnelles.
Un néphron mesure entre 2 et 5 cm. Son glomérule
est destiné à la filtration du sang. Ses
segments tubulaires sont destinés à la réabsorption
et à la sécrétion; on distingue parmi
eux le tube proximal, le segment grêle et le tube distal
qui débouche avec plusieurs autres dans un tube collecteur.
 Le glomérule,
situé dans le cortex (C), est drainé par le tube
proximal. Celuici, représenté en vert,
possède une partie initiale contournée et une partie
rectiligne qui pénètre dans la médullaire
(M). Cette partie droite se prolonge par le segment grêle,
très étroit, représenté en gris. Il
est suivi à son tour par le tube distal, représenté
en brun; sa partie droite regagne le cortex, entre en contact
avec le glomérule et se poursuit par une partie contournée
qui s'abouche au tube collecteur coloré ici en jaune.
Le glomérule,
situé dans le cortex (C), est drainé par le tube
proximal. Celuici, représenté en vert,
possède une partie initiale contournée et une partie
rectiligne qui pénètre dans la médullaire
(M). Cette partie droite se prolonge par le segment grêle,
très étroit, représenté en gris. Il
est suivi à son tour par le tube distal, représenté
en brun; sa partie droite regagne le cortex, entre en contact
avec le glomérule et se poursuit par une partie contournée
qui s'abouche au tube collecteur coloré ici en jaune.
La portion du néphron
comprise entre la partie contournée du tube proximal et
la partie contournée du tube distal est appelée
anse de Henlé. Dans celleci, la séquence
des segments est toujours la même: portion droite du tube
proximal, segment grêle et portion droite du tube distal.
La longueur respective des segments varie suivant la localisation
des glomérules dans le cortex.
Les néphrons internes qui représentent environ
12% de la population totale des néphrons ont une anse très
longue. Elle est constituée essentiellement par le segment
grêle.
Les néphrons externes ont une anse de Henlé
beaucoup plus courte. La branche descendante est formée
par la partie droite du tube proximal et par un court segment
grêle; la courbure de l'anse et la branche ascendante sont
formées par la partie droite du tube distal. Les néphrons
situés dans la partie moyenne du cortex ont une anse de
type intermédiaire.
Précisons cependant que certains, définissant la
anse de Henlé d'un point de vue physiologique, n'y incluent
pas la partie droite du tube proximal.
Les tubes distaux de plusieurs néphrons sont drainés
par un canal collecteur, représenté en jaune. Il
est appelé canal de Bellini à proximité
de son point d'abouchement dans la papille rénale.
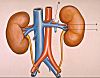 Emanant de l'aorte abdominale, chaque artère rénale
(A) vascularise un rein et se divise au niveau du hile en deux
troncs, pyélique (B) et rétropyélique
(C). Ceuxci se subdivisent dans le sinus en artères
segmentaires dont chacune irrigue une partie du rein, appelée
segment. La division du rein en segments, sur base de la vascularisation,
est surtout utilisée en chirurgie. Le rein peut être
amputé d'un de ses segments sans préjudice pour
le fonctionnement des autres.
Emanant de l'aorte abdominale, chaque artère rénale
(A) vascularise un rein et se divise au niveau du hile en deux
troncs, pyélique (B) et rétropyélique
(C). Ceuxci se subdivisent dans le sinus en artères
segmentaires dont chacune irrigue une partie du rein, appelée
segment. La division du rein en segments, sur base de la vascularisation,
est surtout utilisée en chirurgie. Le rein peut être
amputé d'un de ses segments sans préjudice pour
le fonctionnement des autres.
Les artères segmentaires
se scindent en artères interlobaires (1) qui s'enfoncent
dans le parenchyme au niveau des colonnes de Bertin. Lorsqu'elles
atteignent la base des pyramides, ces artères se divisent
en deux branches qui suivent un trajet parallèle à
la surface de l'organe et portent dès lors le nom d'artères
arquées ou arciformes de premier ordre. Des ramifications
parallèles s'en détachent, ce sont les artères
subarciformes ou arciformes de 2e et 3e ordre (2). Cellesci
donnent à leur tour des branches ascendantes, les artères
interlobulaires (3), perpendiculaires à la surface
du cortex. Elles atteignent la partie la plus externe du cortex,
dépourvue de glomérules, et s'y résolvent
en un réseau capillaire, le plexus souscapsulaire
(4). Certaines cependant se dirigent vers la capsule qu'elles
irriguent; ce sont les artères perforantes de Hyrtl
(5). Les artères arciformes sont anastomosées entre
elles mais la vascularisation corticale, dépendante des
artères interlobulaires, est terminale; l'obstruction d'une
artère ne peut donc être compensée par une
artère voisine. La médullaire quant à elle,
est irriguée par les artères droites (6)
qui se détachent soit des artères arciformes, soit
des artères interlobulaires.
Les artères pyéliques
sont des branches de l'artère rénale. Elles sont
situées dans le sinus du rein et s'y subdivisent en artères
segmentaires. Comme toutes les autres artères elles possèdent
une intima, une média et une adventice.
L'artère interlobaire,
branche de la segmentaire, s'insinue entre deux pyramides de Malpighi.
Dans cette préparation à faible grossissement, elle
est coupée longitudinalement et accompagnée d'une
veine.
Les artères arciformes,
branches des interlobaires, sont beaucoup plus petites et situées
à la jonction corticomédullaire.
Les artères interlobulaires,
branches des arciformes, sont situées dans le cortex. Ce
sont de très petites artères dont la média
est limitée à une ou deux couches musculaires.
L'artère interlobulaire donne naissance à de nombreuses
branches qui pénètrent dans les glomérules
et sont, pour cette raison, appelées artérioles afférentes.
De rares artérioles afférentes naissent cependant
des artères arciformes.
La caractéristique de la vascularisation dans le glomérule
est son organisation en réseau admirable.
Dans un tel système, l'artériole afférente
(1) se résout en une touffe de capillaires, le flocculus
(2), drainé luimême par une artériole
efférente (3), et non par un réseau veineux.
Les deux artérioles sont unies également par l'artère
de Ludwig (4), située en dehors du flocculus et qui
permet de courtcircuiter le système admirable.
Entre l'artère interlobulaire et le glomérule, la
paroi de l'artériole afférente est composée
d'une intima et d'une couche unique de cellules musculaires lisses.
A son entrée dans le glomérule, la paroi de l'artériole afférente est
modifiée; la lame élastique interne qui limite l'intima
disparaît et les cellules musculaires de sa média
entrent dans la constitution de l'appareil juxtaglomérulaire
dont nous parlerons plus loin. Le calibre de l'artériole
efférente est plus petit que celui de l'artériole
afférente, ce qui maintient le gradient de pression nécessaire
à la filtration. Sa structure est, dès la sortie
du glomérule, celle de toute autre artériole.
Lorsque l'artériole efférente
provient des glomérules de Malpighi les plus externes,
elle est de fin calibre et se termine en un réseau capillaire
cortical. Celle des glomérules juxtamédullaires
est plus large et se dirige vers la médullaire où
elle se résout en faisceaux de capillaires à parois
minces et de plus gros calibre qu'habituellement, les vasa
recta, qui engainent à contresens les anses de
Henlé. Nous reparlerons de ces rapports en étudiant
l'histophysiologie des tubes rénaux.
Les veines du rein
suivent le trajet des artères. Les capillaires de la capsule
et du réseau sous-capsulaire se jettent dans des veinules
qui forment les étoiles de Verheyen (1) à
la surface du rein. Le centre de ces étoiles est drainé
par un vaisseau qui s'enfonce dans le cortex et devient la veine
interlobulaire (2) qui draine également le plexus cortical.
Les veines interlobulaires aboutissent aux veines arciformes
(3) qui ellesmêmes confluent pour former les veines
interlobaires (4). Cellesci finalement se rassemblent
pour former les veines pyéliques dans le sinus rénal.
Les vaisseaux lymphatiques sont relativement peu importants.
Ceux de la capsule sont drainés, via la circulation lymphatique
des organes voisins, vers les ganglions aortiques supérieurs.
Ceux du parenchyme forment un réseau entre les tubes, surtout
dans le cortex et confluent en vaisseaux parallèles aux
vaisseaux sanguins. Il n'existe pas de lymphatiques dans les glomérules
et dans les pyramides de Ferrein. L'innervation rénale
dépend du plexus rénal, situé le long de
l'artère rénale depuis son émergence de l'aorte
jusqu'au hile. Il est composé essentiellement de fibres
orthosympathiques issues des ganglions de la chaîne prévertébrale
situés au niveau des 10e, 11e et 12e racines dorsales et
de la 1e lombaire et de quelques fibres parasympathiques en provenance
des nerfs vagues. A partir du plexus rénal, les fibres
nerveuses suivent le trajet des artères jusqu'aux glomérules.
Elles innervent les vaisseaux et l'épithélium des
tubes contournés.
La prochaine leçon sera consacrée à l'étude détaillée du néphron.
 De teinte rougebrun et de consistance ferme, les reins
ont la forme d'un haricot. La face antérieure, visible
sur cette diapositive, est bombée et la face postérieure
est plane. Le bord externe est convexe, et l'interne est échancré.
A ce niveau se trouve le hile, comblé d'un tissu
graisseux de teinte jaunâtre et par où passent les
vaisseaux, les nerfs et l'uretère. L'extrémité
supérieure est plus large que l'inférieure. Chez
l'adulte, les reins mesurent environ 12 cm de haut, 6 cm de large
et 3 cm en épaisseur. Le poids individuel des reins peut
varier, mais le poids global est généralement voisin
de 300 g.
De teinte rougebrun et de consistance ferme, les reins
ont la forme d'un haricot. La face antérieure, visible
sur cette diapositive, est bombée et la face postérieure
est plane. Le bord externe est convexe, et l'interne est échancré.
A ce niveau se trouve le hile, comblé d'un tissu
graisseux de teinte jaunâtre et par où passent les
vaisseaux, les nerfs et l'uretère. L'extrémité
supérieure est plus large que l'inférieure. Chez
l'adulte, les reins mesurent environ 12 cm de haut, 6 cm de large
et 3 cm en épaisseur. Le poids individuel des reins peut
varier, mais le poids global est généralement voisin
de 300 g.

