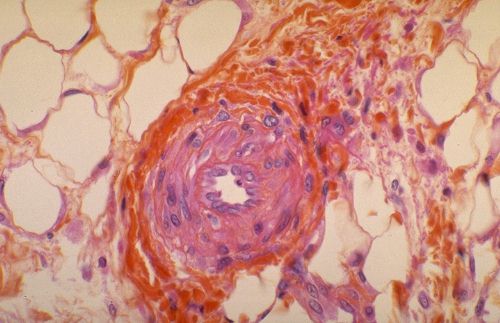
2.6.3. Dépôt d'une lamelle osseuse
La formation d'une lamelle se fait en deux étapes. La
première est l'ostéogénèse. La seconde
est la calcification.
2.6.3.1. Ostéogenèse
L'ostéogénèse est le dépôt
de la phase organique par les ostéoblastes sur un support.
Le support est un tissu mésenchymateux dans l'ossification
membraneuse, une travée cartilagineuse dans l'ossification
endochondrale ou l'os lui-même au cours du remaniement.
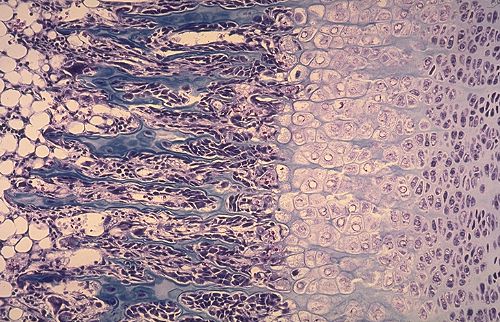
Ossification membraneuse
 Les os du crâne sont élaborés dans du tissu
mésenchymateux. Il s'agit donc d'ossification membraneuse.
La substance fondamentale du tissu conjonctif se modifie et forme
un réseau de travées denses, intensément
colorées en bleu. Des ostéoblastes s'appliquent
contre ces travées et élaborent des lamelles osseuses.
Dès que la lamelle est déposée, le support
n'est plus reconnaissable.
Les os du crâne sont élaborés dans du tissu
mésenchymateux. Il s'agit donc d'ossification membraneuse.
La substance fondamentale du tissu conjonctif se modifie et forme
un réseau de travées denses, intensément
colorées en bleu. Des ostéoblastes s'appliquent
contre ces travées et élaborent des lamelles osseuses.
Dès que la lamelle est déposée, le support
n'est plus reconnaissable.
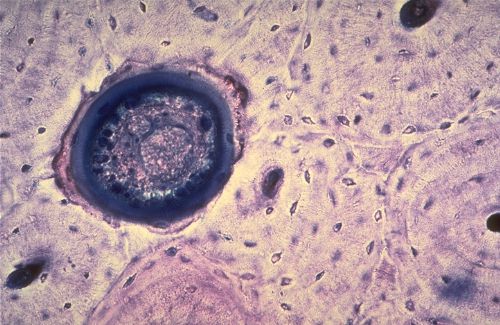
Ossification endochondrale
 Le cartilage hyalin peut devenir le support d'une ossification
endochondrale au cours de l'élaboration et de la croissance
d'un os long. Les chondrocytes, à droite dans la préparation,
s'hypertrophient, sécrètent des phosphatases alcalines
qui déclenchent la calcification de la matrice extracellulaire,
et dégénèrent. Ceci aboutit à la formation
de logettes vides séparées par des travées
calcifiées. Ces logettes sont envahies par des vaisseaux
accompagnés de cellules mésenchymateuses qui prennent
appui sur les travées calcifiées et se transforment
en ostéoblastes.
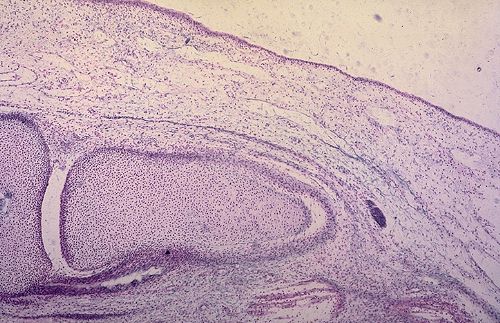
Le cartilage hyalin peut devenir le support d'une ossification
endochondrale au cours de l'élaboration et de la croissance
d'un os long. Les chondrocytes, à droite dans la préparation,
s'hypertrophient, sécrètent des phosphatases alcalines
qui déclenchent la calcification de la matrice extracellulaire,
et dégénèrent. Ceci aboutit à la formation
de logettes vides séparées par des travées
calcifiées. Ces logettes sont envahies par des vaisseaux
accompagnés de cellules mésenchymateuses qui prennent
appui sur les travées calcifiées et se transforment
en ostéoblastes.
Ce support endochondral est représenté ici à
plus fort grossissement. Les travées cartilagineuses sont
colorées en bleu clair et sont dépourvues de cellules.
Les ostéoblastes, disposés en rangées, sont
appliqués contre ces travées. Ce type de support
reste visible, même après le dépôt de
plusieurs lamelles osseuses. Il sera détruit lorsque le
tissu osseux sera remanié.
Ossification sur support osseux
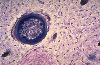 Lors du remaniement de l'os compact, le support d'une nouvelle
ossification est l'os lui-même. Dans cette cavité
osseuse, les cellules mésenchymateuses se sont alignées
le long de la paroi et sont devenues des ostéoblastes.
Lors du remaniement de l'os compact, le support d'une nouvelle
ossification est l'os lui-même. Dans cette cavité
osseuse, les cellules mésenchymateuses se sont alignées
le long de la paroi et sont devenues des ostéoblastes.
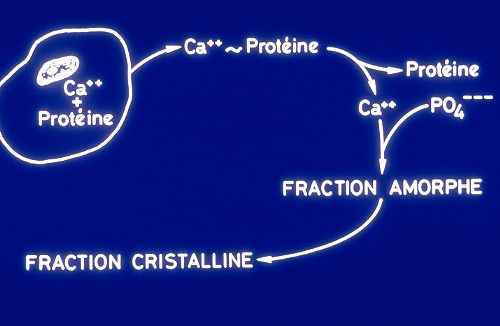
2.6.3.2. Calcification
 La calcification, ou dépôt de la phase minérale,
est la deuxième étape dans la formation d'une lamelle
osseuse. Elle donne à l'os sa consistance. Nous avons vu
que la phase minérale se compose de deux fractions. Une
fraction amorphe constituée de phosphates calciques et
une fraction cristalline constituée d'hydroxyapatites.
Les deux fractions sont produites de façon différente.
Lorsque les ions calcium de la fraction amorphe sont excrétés
par les ostéoblastes ou les ostéocytes, ils sont
liés à une protéine. Ils s'en détachent
immédiatement après l'excrétion. Le calcium
diffuse et précipite sous forme de sels insolubles avec
les ions phosphates de la substance fondamentale. Au fur et à
mesure de sa formation, cette fraction amorphe, très instable,
se transforme en fraction cristalline.
La calcification, ou dépôt de la phase minérale,
est la deuxième étape dans la formation d'une lamelle
osseuse. Elle donne à l'os sa consistance. Nous avons vu
que la phase minérale se compose de deux fractions. Une
fraction amorphe constituée de phosphates calciques et
une fraction cristalline constituée d'hydroxyapatites.
Les deux fractions sont produites de façon différente.
Lorsque les ions calcium de la fraction amorphe sont excrétés
par les ostéoblastes ou les ostéocytes, ils sont
liés à une protéine. Ils s'en détachent
immédiatement après l'excrétion. Le calcium
diffuse et précipite sous forme de sels insolubles avec
les ions phosphates de la substance fondamentale. Au fur et à
mesure de sa formation, cette fraction amorphe, très instable,
se transforme en fraction cristalline.
La formation des cristaux d'hydroxyapatite se fait en deux étapes.
La première étape ou nucléation est
l'apparition des premières mailles du réseau cristallin.
Elle se fait par épitaxie. Ceci signifie que la formation
de la maille est induite et orientée par un modèle
dont l'organisation est adaptée à la configuration
ionique du cristal. Grâce à sa propre configuration,
le modèle peut fixer les ions dans une position identique
à celle qu'ils occupent dans un réseau cristallin.
Dès qu'ils sont dans cette position, les ions se réarrangent
entre eux et forment une maille cristalline. L'identité
du modèle est très discutée.
La deuxième étape de la calcification est l'accrétion.
Au cours de cette phase, les cristaux s'accroissent à partir
des premières mailles produites lors de la nucléation.
Ces mailles peuvent, sur chacune de leurs faces, induire la formation
d'autres mailles rigoureusement identiques qui, à leur
tour, en induisent de nouvelles. Ces inductions en chaîne
provoquent une précipitation massive et brusque de sels
minéraux. Elle atteint d'emblée 75% environ de la
charge minérale totale mais, lorsque les cristaux atteignent
une taille suffisante, le dépôt se ralentit considérablement.
Il faudra plusieurs semaines pour que la minéralisation
soit complète.
L'élaboration d'une lamelle osseuse se fait donc en
deux étapes. Au cours de la première, les ostéoblastes
appliqués contre un support produisent la matrice organique
ou substance ostéoïde et deviennent des ostéocytes
: c'est l'ostéogénèse.
Au cours de la seconde étape, la matrice se charge de
sels minéraux : c'est la calcification. Les sels calciques sont d'abord
déposés sous une forme amorphe. Celle-ci progressivement
devient cristalline. La cristallisation commence par la nucléation
ou dépôt des premières mailles; elle est suivie
de l'accrétion, chaque maille déposée induisant
la formation des autres.
2.6.4. Structure histologique du tissu osseux
 La microradiographie est obtenue en déposant la coupe sur
un film radiographique et en la soumettant aux rayons X. Elle
permet d'apprécier la distribution des sels calciques dans
le tissu. Tout ce qui n'est pas calcifié, y compris les
cavités, est noir.
La microradiographie est obtenue en déposant la coupe sur
un film radiographique et en la soumettant aux rayons X. Elle
permet d'apprécier la distribution des sels calciques dans
le tissu. Tout ce qui n'est pas calcifié, y compris les
cavités, est noir.
2.6.4.1. Type d'os
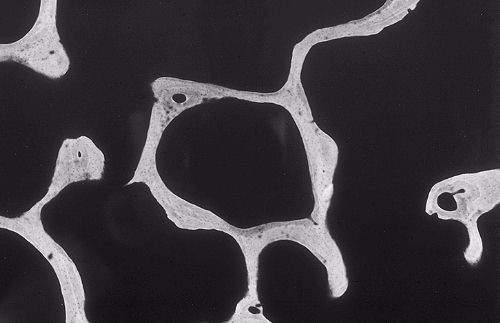
L'os spongieux (vu ici en microradiograhie) est constitué
de travées disposées en réseau autour de
larges espaces irréguliers. Les différentes travées
sont percées de petits orifices noirs qui sont les logettes
des ostéocytes. A plusieurs endroits, le bord des travées
est irrégulier, ce sont des zones de résorption.
En effet, ces travées sont continuellement renouvelées
par le jeu de l'ostéoclasie et de l'ostéogénèse.
L'examen de travées d'os spongieux vu en lumière
polarisée démontre qu'elles sont formées
de lamelles parallèles, alternativement claires et sombres
parce que l'orientation des fibres collagènes change d'une
lamelle à l'autre.
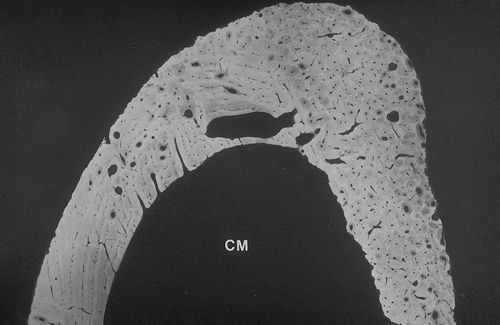
 L'os compact a un aspect très différent.
Nous avons vu qu'il paraît monolithique à l'oeil
nu. Voici la microradiographie d'un segment de diaphyse osseuse
qui révèle la structure complexe de cet os. La cavité
médullaire (CM) est dans le bas de l'image. L'examen à
plus fort grossissement permettra d'expliquer l'aspect hétérogène
des deux types d'os compact : l'os compact primaire et l'os compact
secondaire.
L'os compact a un aspect très différent.
Nous avons vu qu'il paraît monolithique à l'oeil
nu. Voici la microradiographie d'un segment de diaphyse osseuse
qui révèle la structure complexe de cet os. La cavité
médullaire (CM) est dans le bas de l'image. L'examen à
plus fort grossissement permettra d'expliquer l'aspect hétérogène
des deux types d'os compact : l'os compact primaire et l'os compact
secondaire.
L'os primaire est formé de lamelles parallèles
élaborées par appositions successives à partir
du périoste et de l'endoste. Chaque lamelle contient de
nombreux ostéocytes dont les logettes apparaissent comme
de petits points noirs sur cette microradiograhie. Les lignes
noires plus épaisses et discontinues, parallèles
les unes aux autres, sont des canaux vasculaires. Ils sont souvent
réunis entre eux par des canaux transversaux dont les plus
volumineux communiquent avec la cavité médullaire
ou le périoste.
L'os primaire forme la presque totalité de l'os compact
chez le nouveau-né. Au cours de la croissance, il est en
grande partie remplacé par de l'os compact secondaire.
Ceci est la microradiographie d'un segment diaphysaire plus âgé
que le précédent. La cavité médullaire,
dans le haut de l'image, est toujours bordée de lamelles
osseuses parallèles. Le reste de la préparation
est occupé par un grand nombre d'orifices ronds qui sont
la coupe de canaux vasculaires. Tous, ou presque, sont entourés
de leur propre système lamellaire. L'ensemble d'un canal
et des lamelles qui l'entourent est un système de Havers
ou ostéone. Entre les ostéones se trouvent les restes
de l'os primaire.
2.6.4.2. Remaniement haversien
La transformation d'os primaire en os secondaire est le résultat
d'un remaniement continuel. Cette coupe révèle
les différentes étapes de ce remaniement appelé
remaniement haversien. La cavité médullaire est
le large espace clair qui longe le bord gauche de l'image. Tout
le champ est occupé par des ostéones. La largeur
de leur canal est variable. La cavité située en
haut et à gauche, est large et irrégulière,
c'est une cavité de résorption. Répartis
dans la coupe, plusieurs ostéones dont le canal est toujours
entouré d'une couche bleue sont en voie de formation. Les
autres ne possèdent pas de liseré bleu et leur canal
est étroit. Ce sont des ostéones entièrement
achevés.
Dans la microradiographie de la préparation précédente,
la cavité de résorption est un trou noir aux bords
irréguliers. Les ostéones en formation ont un canal
béant aux bords réguliers. Leur surface plus grise
montre qu'ils sont beaucoup moins calcifiés que le reste
de l'os. Les ostéones entièrement achevés
ont un petit canal central. La plupart d'entre eux sont pleinement
calcifiés. L'analyse détaillée de chaque
image nous permet de comprendre comment un ostéone se forme
au cours du remaniement haversien.
La cavité de résorption est la coupe d'un
tunnel creusé dans la masse du tissu osseux. Le forage
de ce tunnel est l'oeuvre d'ostéoclastes en provenance
de la cavité médullaire. Les bords irréguliers
de la cavité sont dentelés par leur érosion.
La cavité est occupée par des vaisseaux dont la
paroi est très mince et de nombreuses cellules mésenchymateuses
dont le noyau est coloré en bleu. Celles-ci s'aligneront
contre la paroi pour s'y transformer en ostéoblastes qui
sécrèteront des fibres collagènes et de la
substance fondamentale. Ils s'enfermeront dans leur production
et deviendront des ostéocytes.
Un ostéone débutant possède quelques
lamelles. Sa limite externe, sinueuse, appelée ligne cimentante,
est la trace de l'ancienne cavité de résorption
dans laquelle l'ostéone s'est formé. La lumière
du canal est large, les vaisseaux bien visibles. De nouveaux ostéoblastes
sont appliqués contre la lamelle à peine formée
qui dès lors sert de support à une lamelle plus
interne. Le canal primitivement large se rétrécit
ainsi lamelle par lamelle.
Un ostéone plus avancé est formé de
plusieurs lamelles. Les lamelles périphériques sont
blanches. Elles sont creusées de petites lacunes qui contiennent
les ostéocytes. La lamelle interne est colorée en
bleu. C'est le liseré préosseux, substance ostéoïde
non calcifiée de cet ostéone. La zone limite entre
le liseré préosseux et la première lamelle
calcifiée s'appelle ligne frontière. Le canal est
encore large. Son pourtour est occupé par une couche d'ostéoblastes
teintés en bleu foncé. Ici encore, les ostéoblastes
produiront une nouvelle lamelle ou plus exactement un nouveau
liseré préosseux, tandis que l'actuel se calcifie.
Dans cet ostéone beaucoup plus développé,
le canal central est réduit. La présence d'un liseré
préosseux démontre cependant que le dépôt
des lamelles les plus centrales n'est pas terminé.
 Cet ostéone achevé n'a plus de liseré
préosseux. Il est séparé du tissu voisin
par une ligne sinueuse, la ligne cimentante qui, rappelons-le,
est la limite de la cavité de résorption dans laquelle
il s'est formé. Le canal central ou canal vasculaire de
Havers est étroit. Il contient des vaisseaux qui ne sont
pas visibles dans cette préparation. La masse de l'ostéone
est formée de lamelles successives, disposées concentriquement
par rapport au canal central. Chaque lamelle est creusée
de quelques logettes qui contiennent des ostéocytes. La
fine striation radiaire à travers toute l'épaisseur
de l'ostéone est due aux nombreux canalicules qui relient
les logettes des ostéocytes.
Cet ostéone achevé n'a plus de liseré
préosseux. Il est séparé du tissu voisin
par une ligne sinueuse, la ligne cimentante qui, rappelons-le,
est la limite de la cavité de résorption dans laquelle
il s'est formé. Le canal central ou canal vasculaire de
Havers est étroit. Il contient des vaisseaux qui ne sont
pas visibles dans cette préparation. La masse de l'ostéone
est formée de lamelles successives, disposées concentriquement
par rapport au canal central. Chaque lamelle est creusée
de quelques logettes qui contiennent des ostéocytes. La
fine striation radiaire à travers toute l'épaisseur
de l'ostéone est due aux nombreux canalicules qui relient
les logettes des ostéocytes.
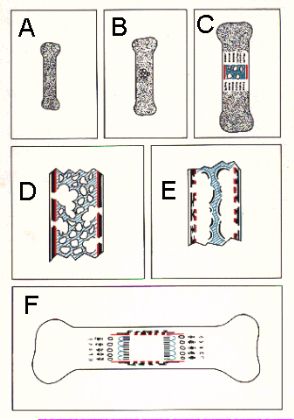
Ce schéma de la formation d'un ostéone permet de
suivre son évolution. Chaque phase est, à droite,
représentée en coupe transversale et correspond
à peu de choses près aux images précédentes.
Le début de l'ostéone est un tunnel de résorption
foré dans la masse osseuse par des ostéoclastes.
Il est représenté en D. Les ostéoclastes
sont suivis d'un vaisseau entouré de cellules mésenchymateuses.
Celles-ci se rangent contre la paroi de la cavité de résorption
et deviennent des ostéoblastes. Ils déposent une
première lamelle osseuse, d'abord non calcifiée,
rétrécissant d'autant le canal central. Ce stade
est représenté en C. D'autres cellules mésenchymateuses
se transforment en nouveaux ostéoblastes qui déposent
de nouvelles lamelles en dedans de la première. Elles apparaissent
en B et en A. Le dépôt continue jusqu'à ce
que le canal initial soit réduit à un pertuis beaucoup
plus étroit : le canal vasculaire de Havers de l'ostéone
définitif. Le remaniement haversien se poursuit durant
toute la vie. Il est très important au cours de la croissance,
ralentit considérablement en fin de croissance et est très
lent au cours de la vie adulte.
Toute pièce osseuse est enveloppée d'une membrane
appelée périoste. Il est composé de
deux couches mal délimitées. La couche externe est
fibreuse. Les faisceaux collagènes, colorés en bleu,
sont parallèles à la surface de l'os. Par endroits,
quelques faisceaux se coudent, traversent la couche interne et
sont incorporés dans l'épaisseur de l'os. Ils servent
à ancrer le périoste. La couche interne est celluleuse.
Ses cellules mésenchymateuses se transforment en ostéoblastes,
s'alignent contre l'os déjà formé et produisent
de nouvelles lamelles osseuses. Cette apposition, au cours de
la croissance, épaissit l'os.
2.6.5. Elaboration et croissance d'un os long
 Nous avons vu que les os plats sont d'origine membraneuse. L'os
long est d'origine endochondrale. Chez l'embryon, il est préfiguré
par un modèle cartilagineux. Très tôt
au cours du développement embryonnaire, une pièce
squelettique ne contient donc que du cartilage hyalin; les cellules
mésenchymateuses voisines se tassent autour de lui pour
ébaucher le périchondre qui, par apposition, allonge
et épaissit la pièce squelettique. La pièce
squelettique et son périchondre se trouvent au centre de
l'image et sont entourés de tissu conjonctif muqueux.
Nous avons vu que les os plats sont d'origine membraneuse. L'os
long est d'origine endochondrale. Chez l'embryon, il est préfiguré
par un modèle cartilagineux. Très tôt
au cours du développement embryonnaire, une pièce
squelettique ne contient donc que du cartilage hyalin; les cellules
mésenchymateuses voisines se tassent autour de lui pour
ébaucher le périchondre qui, par apposition, allonge
et épaissit la pièce squelettique. La pièce
squelettique et son périchondre se trouvent au centre de
l'image et sont entourés de tissu conjonctif muqueux.
Après une phase de simple croissance par apposition de
cartilage, les premières modifications apparaissent au
centre de la pièce squelettique. Le cartilage hyalin
devient hypertrophique et se calcifie. Lorsque cette hypertrophie
atteint les parties latérales de la pièce, les cellules
mésenchymateuses du périchondre ne se transforment
plus en chondroblastes mais en ostéoblastes qui élaborent
une première lamelle osseuse, ligne plus colorée
contre le cartilage. Le périchondre est donc, à
cet endroit, devenu périoste.
Très rapidement le manchon osseux s'épaissit. Sans
cesse en effet, les cellules mésenchymateuses en provenance
du périoste se transforment en ostéoblastes qui,
enfermés dans la matrice organique qu'ils produisent, deviennent
des ostéocytes. Ce manchon osseux est coloré en
noir et représente la première ébauche de
la future diaphyse. Les travées du cartilage hypertrophique,
elles aussi calcifiées, sont colorées en bleu mais
dépourvues d'ostéocytes.
Ensuite, des capillaires en provenance du périoste traversent
cette couche osseuse et érodent le cartilage hypertrophique.
En même temps, l'hypertrophie du cartilage s'étend
vers les extrémités, ce qui nécessairement
allonge la diaphyse osseuse.
Voici, dans une étape ultérieure du développement,
le résultat de cette invasion conjonctivo-vasculaire. Les
vaisseaux ont creusé de larges cavités. Les cellules
mésenchymateuses qui accompagnent ces vaisseaux se sont
disposées contre les supports endochondraux calcifiés
puis, utilisant ce support, se sont transformées en ostéoblastes
et ont produit des travées osseuses. Celles-ci ont été
presque immédiatement détruites par des ostéoclastes
qui creusent ainsi la cavité médullaire.
La première partie du développement d'un os long,
celle qui aboutit à la formation de sa diaphyse peut se
résumer en quelques schémas.
 La pièce squelettique est composée d'abord uniquement
de cartilage hyalin (a). Un nodule de cartilage hypertrophique
calcifié apparaît au centre de la pièce (b).
Le cartilage hypertrophique, coloré en bleu, atteint le
manchon périchondral et induit la formation d'une lamelle
osseuse, colorée en rouge (c). L'invasion conjonctivo-vasculaire
provenant du périoste creuse des cavités dans le
cartilage hypertrophique. Le périoste dépose de
nouvelles lamelles osseuses, colorées en noir, sur la face
externe de la première (d). Des ostéoblastes déposent
des lamelles osseuses sur les travées cartilagineuses (e).
Des ostéoclastes détruisent les travées endochondrales
centrales (f). Ainsi se forme la cavité médullaire.
La pièce squelettique est composée d'abord uniquement
de cartilage hyalin (a). Un nodule de cartilage hypertrophique
calcifié apparaît au centre de la pièce (b).
Le cartilage hypertrophique, coloré en bleu, atteint le
manchon périchondral et induit la formation d'une lamelle
osseuse, colorée en rouge (c). L'invasion conjonctivo-vasculaire
provenant du périoste creuse des cavités dans le
cartilage hypertrophique. Le périoste dépose de
nouvelles lamelles osseuses, colorées en noir, sur la face
externe de la première (d). Des ostéoblastes déposent
des lamelles osseuses sur les travées cartilagineuses (e).
Des ostéoclastes détruisent les travées endochondrales
centrales (f). Ainsi se forme la cavité médullaire.
Au terme de cette première transformation, l'os long
est composé de ses différents segments : diaphyse,
épiphyses et métaphyses.
La diaphyse est un cylindre d'os compact autour de la cavité
médullaire; elle est enveloppée de périoste.
L'épiphyse est d'abord un nodule de cartilage hyalin. Assez
tardivement par rapport à la diaphyse, les cellules centrales
deviennent hypertrophiques. Cette zone d'hypertrophie croît
de façon radiaire et, par ses transformations successives,
sert de support au dépôt d'os spongieux. En même
temps le volume de l'épiphyse augmente par apposition de
cartilage. Celle-ci s'arrête lorsque l'hypertrophie du cartilage
atteint le périchondre. Le périchondre devient alors
périoste qui enveloppe l'épiphyse d'une coque d'os
compact. Seule la région articulaire, dépourvue
de périchondre, reste cartilagineuse.
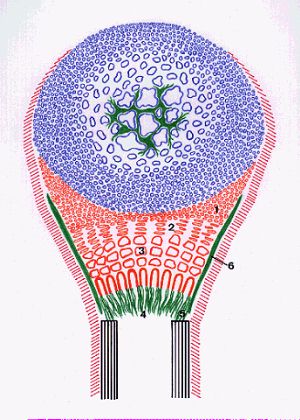
 La métaphyse est située entre la diaphyse et l'épiphyse.
Elle contient le cartilage de conjugaison composé d'une
couche de cartilage hyalin (1), d'une couche de cartilage sérié
(2), d'une couche de cartilage hypertrophique (3) et d'une zone
d'invasion conjonctivo-vasculaire où se forment des travées
osseuses centrales (4) et des travées osseuses périphériques
(5). Celles-ci dessinent un cône tronqué d'os spongieux
dont la petite base s'appuie sur le cylindre diaphysaire et dont
la grande base est en continuité avec la périphérie
du cartilage de conjugaison. Le cartilage hyalin est en rapport
avec le périchondre. Les autres régions sont séparées
du périoste par une fine lame osseuse (6).
La métaphyse est située entre la diaphyse et l'épiphyse.
Elle contient le cartilage de conjugaison composé d'une
couche de cartilage hyalin (1), d'une couche de cartilage sérié
(2), d'une couche de cartilage hypertrophique (3) et d'une zone
d'invasion conjonctivo-vasculaire où se forment des travées
osseuses centrales (4) et des travées osseuses périphériques
(5). Celles-ci dessinent un cône tronqué d'os spongieux
dont la petite base s'appuie sur le cylindre diaphysaire et dont
la grande base est en continuité avec la périphérie
du cartilage de conjugaison. Le cartilage hyalin est en rapport
avec le périchondre. Les autres régions sont séparées
du périoste par une fine lame osseuse (6).
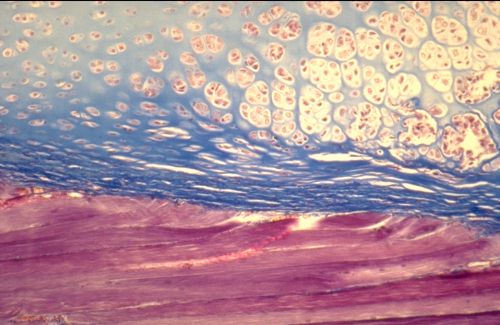
Voici un segment du cartilage de conjugaison métaphysaire.
La couche proche des épiphyses, en haut dans l'image, est
hyaline. La couche moyenne, au centre de l'image, est le cartilage
sérié formé de longs groupes isogéniques
axiaux. Leur élaboration est due aux mitoses des chondrocytes
de la partie inférieure de la zone hyaline. Sous le cartilage
sérié se trouvent le cartilage hypertrophique, puis
la zone d'invasion conjonctivo-vasculaire.
Le cartilage sérié est formé de l'empilement
de chondrocytes aplatis. La cellule du cartilage hyalin ajoute
par des mitoses successives de nouveaux éléments
à l'extrémité proximale de ce groupe isogénique
axial. Chaque groupe est isolé de ses voisins par une travée
de matrice organique. A l'extrémité distale du groupe,
le chondrocyte devient hypertrophique.
L'aspect du cartilage hypertrophique est dû à l'accroissement
du volume cellulaire et donc des logettes. Dès le début
de son accroissement, figuré en haut, le chondrocyte sécrète
de la phosphatase alcaline. Ensuite, il dégénère.
Son cytoplasme devient vacuolaire. Son noyau entre en caryolyse.
A la fin de cette dégénérescence, en bas
dans l'image, il ne reste dans la logette que des débris
cellulaires. A cet endroit, situé à la limite de
la cavité médullaire, les logettes vides des différents
groupes isogéniques sont séparées par d'épaisses
travées longitudinales calcifiées. Les vaisseaux
en provenance de la cavité médullaire détruisent
les fines cloisons transversales qui séparent les logettes
vides d'un groupe donné, envahissent ces logettes et se
trouvent ainsi dans l'axe des groupes isogéniques.
La plupart des transformations de l'os long au cours de sa croissance
siègent dans la métaphyse. Le cartilage disparaît
progressivement dans sa partie centrale mais s'accroît en
périphérie. Les travées osseuses centrales
sont résorbées pour maintenir et accroître
la cavité médullaire. Les travées osseuses
périphériques de la métaphyse sont incorporées
par segments dans la diaphyse.
Les modifications du cartilage sont schématisées
ici dans trois phases successives de la croissance. Pourquoi
le cartilage disparaît-il dans sa partie centrale? Chaque
groupe isogénique axial est alimenté par les divisions
successives d'une cellule du cartilage hyalin. Dès que
le pouvoir mitotique de cette cellule s'épuise, le groupe
n'est plus alimenté. L'hypertrophie cellulaire toujours
suivie de dégénérescence continue cependant
et atteint finalement la couche hyaline. La zone centrale est
la plus rapidement atteinte parce que la plus ancienne. Ainsi
se crée une brèche, rapidement envahie par des vaisseaux,
qui met en communication la cavité médullaire et
les mailles du treillis spongieux épiphysaire.
Comment le cartilage s'accroît-il en périphérie?
Par apposition de nouveaux chondrocytes qui se différencient
aux dépens des cellules mésenchymateuses du périchondre
en regard du cartilage hyalin. Ceux-ci produiront à leur
tour des groupes isogéniques axiaux et donc des travées
endochondrales, situées en dehors des travées les
plus externes du stade précédent.
Comment les travées périphériques sont-elles
incorporées dans la diaphyse lorsqu'elles ont atteint
une longueur suffisante? L'incorporation du segment métaphysaire
dans la diaphyse suppose : la transformation d'un treillis d'os
spongieux en un bloc d'os compact; l'alignement des diamètres
du segment métaphysaire sur les diamètres diaphysaires.
La transformation de l'os spongieux en os compact est due à
une activité ostéogénique intense de l'endoste.
Elle comble toutes les mailles et emprisonne ainsi l'os endochondral
dans une masse compacte irrégulière. L'endoste aligne
aussi le diamètre interne du segment métaphysaire
sur le diamètre interne de la diaphyse en déposant
des lamelles régulières d'os primaire. L'alignement
du diamètre externe du segment métaphysaire et du
diamètre externe de la diaphyse suppose la suppression
d'un excès d'os spongieux. Elle est réalisée
par les ostéoclastes très nombreux dans cette région
du périoste. Le schéma représente cette transformation
en coupes longitudinales et transversales, à deux phases
successives de la croissance. Les travées endochondrales
sont figurées en orange, l'os compact d'origine endostéale
en rouge et l'os résorbé par le périoste
est figuré par les espaces entourés de lignes pointillées.
Dès que le segment métaphysaire est devenu diaphyse,
il subit un remodelage : ces nouvelles modifications sont nécessaires
pour élargir le cylindre diaphysaire et pour le donner
sa forme caractéristique.
L'élargissement de la diaphyse est le résultat d'une
apposition continue d'os primaire par le périoste, représentée
en rouge. La cavité médullaire est élargie
grâce à l'activité ostéoclasique de
l'endoste, représentée en pointillés. L'apposition
périostique et l'ostéoclasie endostéale ne
sont pas identiques sur toute la périphérie. Des
zones d'apposition et de résorption localisées modifient
la forme de la diaphyse. Ce schéma, où les zones
d'apposition sont indiquées en rouge et les zones d'ostéoclasie
en bleu, montre comment ce remodelage peut transformer une région
osseuse primitivement annulaire. Il se superpose à l'épaississement
progressif de la diaphyse.
Vous terminerez votre étude en analysant les quatre dernières
images. De quel(s) tissu(s) s'agit-il et pourquoi?
 Réponse
Réponse
 Réponse
Réponse
 Réponse
Réponse
 Réponse
Réponse
Leçon suivante: Le sang et les systèmes de défense
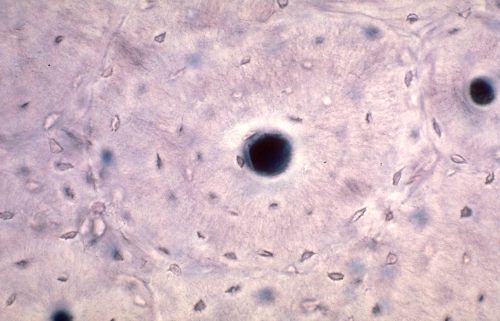
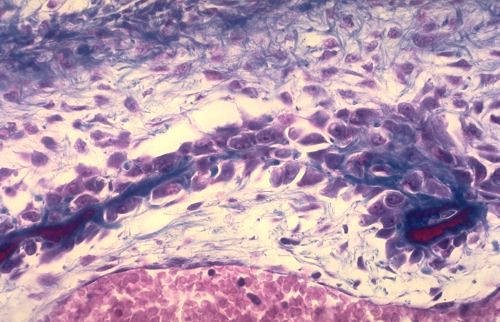 Les os du crâne sont élaborés dans du tissu
mésenchymateux. Il s'agit donc d'ossification membraneuse.
La substance fondamentale du tissu conjonctif se modifie et forme
un réseau de travées denses, intensément
colorées en bleu. Des ostéoblastes s'appliquent
contre ces travées et élaborent des lamelles osseuses.
Dès que la lamelle est déposée, le support
n'est plus reconnaissable.
Les os du crâne sont élaborés dans du tissu
mésenchymateux. Il s'agit donc d'ossification membraneuse.
La substance fondamentale du tissu conjonctif se modifie et forme
un réseau de travées denses, intensément
colorées en bleu. Des ostéoblastes s'appliquent
contre ces travées et élaborent des lamelles osseuses.
Dès que la lamelle est déposée, le support
n'est plus reconnaissable.