2.3. Tissu réticulé
 Les deux composants du tissu
réticulé peuvent être
mis en évidence par l'éosine, qui colore les cellules
en rouge, et par une imprégnation à l'argent, qui
colore en noir les fibres collagènes de type III. Les fibres
dessinent un réseau à larges mailles ouvertes qui
contiennent de petites cellules sanguines rondes. Les cellules
réticulées sont accrochées aux fibres. Elles
sont allongées et leurs contours sont mal définis.
L'une d'entre elles est désignée par une flèche.
Les deux composants du tissu
réticulé peuvent être
mis en évidence par l'éosine, qui colore les cellules
en rouge, et par une imprégnation à l'argent, qui
colore en noir les fibres collagènes de type III. Les fibres
dessinent un réseau à larges mailles ouvertes qui
contiennent de petites cellules sanguines rondes. Les cellules
réticulées sont accrochées aux fibres. Elles
sont allongées et leurs contours sont mal définis.
L'une d'entre elles est désignée par une flèche.
Dans les préparations habituelles, les
cellules réticulées
sont bien visibles, mais les fibres ne le sont pas. Les cellules
réticulées, désignées par des flèches,
sont étoilées et leurs prolongements dessinent un
réseau superposable au réseau de fibres décrit
dans l'image précédente. Deux faisceaux de fibres
collagènes de type I, colorés en bleu, traversent
la préparation.
Le tissu réticulé ne contient donc que deux des
trois constituants habituels des tissus conjonctifs. Il est dépourvu
de substance fondamentale. Les fibres argyrophiles sont formées
de collagène type III et disposées en réseau.
Les cellules réticulées sont accrochées aux
fibres et s'anastomosent par leurs prolongements. Le tissu réticulé
forme le squelette des ganglions lymphoïdes, de la rate et
de la moelle osseuse; ses mailles contiennent soit de la lymphe
soit des cellules sanguines.
2.4. Tissu adipeux
 A l'oeil nu, le tissu adipeux paraît homogène et
nacré; il est, pour cette raison, appelé graisse
blanche. En voici dans la peau; les cellules sont groupées
en lobules séparés par des travées de tissu
conjonctif dense. Leur cytoplasme est presque entièrement
occupé par une volumineuse goutte lipidique qui refoule
le noyau, désigné par une flèche, contre
la membrane plasmique, d'où son nom de graisse uniloculaire.
On ne voit pas les triglycérides, car ils ont été
extraits lors de la déshydratation et de l'enrobage; il
ne reste, à leur emplacement, qu'une plage vide et donc
non colorée. Chaque lobule est desservi par une petite
artère qui se divise en un riche réseau capillaire
intralobulaire. La vascularisation n'est pas visible sur cette
image.
A l'oeil nu, le tissu adipeux paraît homogène et
nacré; il est, pour cette raison, appelé graisse
blanche. En voici dans la peau; les cellules sont groupées
en lobules séparés par des travées de tissu
conjonctif dense. Leur cytoplasme est presque entièrement
occupé par une volumineuse goutte lipidique qui refoule
le noyau, désigné par une flèche, contre
la membrane plasmique, d'où son nom de graisse uniloculaire.
On ne voit pas les triglycérides, car ils ont été
extraits lors de la déshydratation et de l'enrobage; il
ne reste, à leur emplacement, qu'une plage vide et donc
non colorée. Chaque lobule est desservi par une petite
artère qui se divise en un riche réseau capillaire
intralobulaire. La vascularisation n'est pas visible sur cette
image.
Lorsque les adipocytes sont jeunes et ne contiennent que peu de
lipides, ceux-ci sont dispersés dans le cytoplasme sous
forme de petites gouttelettes. Au fur et à mesure que la
quantité de lipides augmente, les gouttelettes confluent
pour finalement donner à la cellule son aspect uniloculaire
caractéristique. La taille des cellules dépend de
la quantité de lipides accumulés. Les vaisseaux
peuvent être identifiés grâce à la présence
de globules rouges, indiqués par des flèches.
Dans le tissu adipeux coloré à l'argent, seules
les fibres reticulées sont mises en évidence. Les
travées de tissu conjonctif dense délimitent cinq
lobules. Dans chacun d'eux, l'emplacement des adipocytes est délimité
par les fibres réticulées.
Le tissu adipeux est un tissu conjonctif où prédominent
des cellules spécialisées dans l'accumulation des
triglycérides, appelées cellules adipeuses ou adipocytes.
Chaque adipocyte est entouré d'un réseau de fibres
collagènes de type III. La graisse blanche est
d'abord un réservoir d'énergie. En effet, les graisses
ingérées lors d'un repas sont stockées sous
forme de triglycérides dans les adipocytes, puis libérées
dans le sang selon les besoins en énergie entre les repas.
Elle permet donc une alimentation intermittente. Elle joue aussi
un rôle d'isolant thermique et de protection mécanique.
Dans la graisse brune, les adipocytes diffèrent
des précédents.
Ils sont plus petits et contiennent de nombreuses gouttelettes
lipidiques qui confluent rarement en une goutte unique. Ce sont
des adipocytes de la graisse multiloculaire, appelée aussi
graisse brune à cause de sa couleur naturelle, due aux
cytochromes des nombreuses mitochondries.
La graisse brune est surtout abondante chez l'embryon.
Elle diminue au cours de la croissance et est réduite à
presque rien chez l'adulte. Les adipocytes de la graisse brune
contiennent de très nombreuses mitochondries capables d'oxyder
les acides gras très rapidement. L'énergie d'oxydation
n'y est pas utilisée pour la synthèse d'ATP, mais
est dégagée sous forme de chaleur. Ceci explique
le rôle de la graisse brune dans l'adaptation au froid chez
le nourrisson et chez les animaux qui hibernent.
2.5. Cartilage
 Le cartilage constitue le squelette définitif de certains
poissons et le squelette provisoire des embryons de mammifères.
Chez ceux-ci, le cartilage est progressivement remplacé
par un squelette osseux. Chez l'adulte, des pièces squelettiques
cartilagineuses persistent dans l'oreille, le nez, le larynx,
la trachée et les bronches. Au niveau des surfaces articulaires,
il empêche, avec le liquide synovial, le frottement des
pièces osseuses l'une contre l'autre. Au niveau du cartilage
de conjugaison, il intervient dans la croissance des os longs.
Seules les pièces cartilagineuses du squelette sont représentées
dans ce schéma.
Le cartilage constitue le squelette définitif de certains
poissons et le squelette provisoire des embryons de mammifères.
Chez ceux-ci, le cartilage est progressivement remplacé
par un squelette osseux. Chez l'adulte, des pièces squelettiques
cartilagineuses persistent dans l'oreille, le nez, le larynx,
la trachée et les bronches. Au niveau des surfaces articulaires,
il empêche, avec le liquide synovial, le frottement des
pièces osseuses l'une contre l'autre. Au niveau du cartilage
de conjugaison, il intervient dans la croissance des os longs.
Seules les pièces cartilagineuses du squelette sont représentées
dans ce schéma.
Les cellules du cartilage ou chondrocytes sont isolées
dans de petites lacunes ou chondroplastes. Elles n'ont pas de
forme particulière, mais épousent celle de leur
cavité. Leur noyau volumineux est rond et possède
un ou deux nucléoles. Leur cytoplasme est granuleux. Les
chondroplastes sont séparés par la matrice organique.
2.5.1. Types de cartilages
 L'aspect de la matrice permet de distinguer trois types de cartilage:
le cartilage hyalin, le cartilage fibreux et le cartilage
élastique.
L'aspect de la matrice permet de distinguer trois types de cartilage:
le cartilage hyalin, le cartilage fibreux et le cartilage
élastique.
Le cartilage hyalin est le plus fréquent; on l'appelle
ainsi parce qu'il est vitreux à l'état frais. Sa
matrice paraît homogène en microscopie optique parce
que la partie fibrillaire est faite d'un feutrage de petites fibres
collagènes de type II, responsables de la résistance
du cartilage. Les glycosaminoglycanes de la substance fondamentale
sont particulièrement riches en radicaux acides, ce qui
explique la métachromasie du cartilage. Ces radicaux sont
très hydrophiles, ce qui explique la haute teneur en eau
et donc l'élasticité du cartilage.
Dans le cartilage fibreux, la composante fibrillaire est faite
d'épais faisceaux de fibres collagènes de type I,
donc bien visibles en microscopie optique. Ce cartilage ressemble
donc à un tissu conjonctif dense; il n'en diffère
que par ses chondrocytes, très différents de fibroblastes.
Dans le cartilage élastique, propre à certains ligaments,
ces faisceaux fibrillaires sont remplacés par des lames
élastiques.
Toute pièce cartilagineuse, sauf le cartilage articulaire,
est enveloppé d'une membrane, le périchondre, qui
n'est bien développé que dans les pièces
cartilagineuses en croissance. Il se trouve dans la partie supérieure
de l'image; on lui décrit habituellement deux couches,
mal délimitées l'une de l'autre. L'externe est plus
riche en fibres collagènes. L'interne est riche en cellules
mésenchymateuses.
Autour d'une pièce cartilagineuse qui n'est pas en croissance,
le périchondre est fin et souvent réduit à
une capsule conjonctive très mince.
2.5.2. Modes de croissance du cartilage
La croissance du cartilage peut se faire de deux façons.
Elle peut se faire en périphérie à partir
des cellules mésenchymateuses du périchondre ou
être interstitielle et se fait alors à partir des
mitoses de chondrocytes.
Dans la croissance périphérique (A), le cartilage
s'épaissit par apposition de couches externes issues du
périchondre. Les cellules mésenchymateuses de la
couche interne du périchondre se transforment en chondroblastes
qui sécrètent de la matrice et s'enferment ainsi
dans une logette. Dans la croissance interstitielle du
cartilage (B), un chondrocyte se divise plusieurs fois de suite
dans sa logette et, après chaque division, la cellule-fille
sécrète de la matrice et s'éloigne ainsi
de la cellule-mère. Les divisions peuvent se faire dans
tous les sens et produisent alors un groupe isogénique
coronaire. Elles peuvent se faire dans un sens déterminé
et donnent naissance à un groupe isogénique axial.
Les images suivantes sont des exemples de ces types de croissance.
Au cours de l'apposition, les cellules mésenchymateuses
internes du périchondre se transforment progressivement
en chondrocytes.
Le groupe isogénique coronaire est formé
dans le cartilage par multiplications successives d'un chondrocyte
sans orientation déterminée.
Le groupe isogénique axial est également
formé dans le cartilage par les divisions successives d'une
cellule. Elles se réalisent de telle façon que les
cellules-filles sont empilées les unes sur les autres.
Le cartilage peut subir de nombreuses modifications. Certaines
sont anormales; l'apparition de fissures ou de structures fibrillaires
inhabituelles sont des signes pathologiques de vieillissement
ou de dégénérescence. D'autres sont physiologiques;
la matrice du cartilage peut se calcifier lorsqu'elle doit servir
de support à l'ossification ou lorsqu'elle donne insertion
à un ligament.
Le cartilage est donc un tissu conjonctif entrant dans la composition
du squelette. Il est formé de chondrocytes isolés
dans une logette et d'une matrice organique importante dont la
composante fibrillaire détermine le type tissulaire. Sa
croissance se fait à partir du périchondre ou est
interstitielle.
2.6. Le tissu osseux
2.6.1. Aspects macroscopiques
 Le tissu osseux est un tissu conjonctif calcifié à
structure lamellaire. C'est le composant le plus important
du squelette. Il a donc avant tout un rôle de soutien et
de protection, mais il intervient aussi dans le métabolisme
des sels minéraux.
Le tissu osseux est un tissu conjonctif calcifié à
structure lamellaire. C'est le composant le plus important
du squelette. Il a donc avant tout un rôle de soutien et
de protection, mais il intervient aussi dans le métabolisme
des sels minéraux.
Le tissu osseux existe sous deux formes. Lorsqu'il paraît
dense à l'oeil nu et dépourvu de cavités,
on l'appelle "os compact". Lorsqu'au contraire,
il est formé de minces travées disposées
en réseau autour de nombreuses cavités, il a l'aspect
d'une éponge et s'appelle "os spongieux".
Dans cette coupe macroscopique d'un os long, l'os compact délimite
entre autres la cavité de la partie droite du cliché;
la partie supérieure contient surtout du tissu osseux spongieux.
Anatomiquement, on distingue trois types de pièces
osseuses: l'os plat, l'os court et l'os long. L'os plat est formé
de deux tables d'os compact séparées par une couche
d'os spongieux, la diploé. L'os court, dont toutes les
dimensions sont à peu près égales, est formé
d'une masse spongieuse entourée d'une fine gaine d'os compact.
L'os long comprend plusieurs parties. Ses deux extrémités
ou épiphyses sont formées d'os spongieux entouré
d'une lame d'os compact. La partie moyenne ou diaphyse est un
cylindre d'os compact creusé d'une large cavité
centrale, la cavité médullaire. Entre la partie
moyenne et chaque extrémité se trouve une zone de
transition, la métaphyse, occupée par le cartilage
de conjugaison au cours de la croissance. Toute pièce osseuse
est recouverte d'une membrane, le périoste. Les cavités
internes sont tapissées par l'endoste, fine lame mésenchymateuse
qui n'est pas visible à l'oeil nu.
2.6.2. Lamelle osseuse
 Quelle que soit sa forme macroscopique, le tissu osseux est formé
de lamelles osseuses dont la disposition varie selon le type d'os.
L'épaisseur de ces lamelles peut atteindre 7 µm. Elles
sont parallèles et étroitement accollées
les unes aux autres. Elles contiennent de petites cavités
qui logent, chacune, un ostéocyte et sont reliées
entre elles par de fins canalicules. Cette structure lamellaire
est bien visible sur des coupes épaisses obtenues par usure.
Quelle que soit sa forme macroscopique, le tissu osseux est formé
de lamelles osseuses dont la disposition varie selon le type d'os.
L'épaisseur de ces lamelles peut atteindre 7 µm. Elles
sont parallèles et étroitement accollées
les unes aux autres. Elles contiennent de petites cavités
qui logent, chacune, un ostéocyte et sont reliées
entre elles par de fins canalicules. Cette structure lamellaire
est bien visible sur des coupes épaisses obtenues par usure.
2.6.2.1. Matrice extracellulaire
La lamelle osseuse, l'unité du tissu osseux, est formée
d'une matrice extracellulaire, comprenant une phase organique
et une phase minérale, et de cellules. La phase organique
est composée pour 90% de fibres collagènes et pour
environ 10% de substance fondamentale. Celle-ci contient surtout
de l'acide hyaluronique et des glycosaminoglycanes non associés
en protéoglycanes. Sa teneur en eau, d'environ
25%, est très faible. Les fibres collagènes sont
du type I et du type V. Toutes les fibres ont, dans leur lamelle,
la même orientation, mais cette orientation change de lamelle
en lamelle. Cette disposition des fibres collagènes est
responsable de l'alternance de lamelles isotropes sombres et anisotropes
claires dans un tissu osseux examiné en lumière
polarisée.
La phase minérale est composée d'une fraction
amorphe et d'une fraction cristalline. La fraction amorphe
contient divers phosphates calciques. La fraction cristalline,
la plus importante, est constituée de cristaux d'hydroxyapatite.
Le cristal d'hydroxyapatite, représenté en A, a
l'aspect d'une longue aiguille. C'est un solide dont les atomes
sont organisés en ensembles répétés
régulièrement dans les trois dimensions de l'espace.
L'unité du cristal est la maille; elle est représentée
en C. La maille est un parallélipipède qui se définit
par trois axes et trois angles. Trois mailles s'agencent en formant
un prisme droit à base hexagonale. Ces prismes sont représentés
à fort grossissement en B. Une série de prismes
s'associe en un cristal. Les cristaux sont tassés les uns
contre les autres, autour des fibres collagènes. La partie
organique est donc entièrement emprisonnée dans
la masse cristalline.
2.6.2.2. Cellules
Il existe dans le tissu osseux trois types de cellules :
Ostéoblaste
 L'ostéoblaste est la cellule qui construit la lamelle
osseuse. Nous verrons plus loin que toute ostéogénèse
se fait contre un support. Le support de l'ostéogénèse
est représenté ici par une bande longitudinale colorée
en bleu. Les ostéoblastes se disposent contre ce support
en une couche continue de cellules cubiques. Leur cytoplasme est
intensément basophile, ce qui se traduit dans cette préparation
par une couleur violette. Cette basophilie est due à la
richesse en réticulum endoplasmique rugueux. Le noyau rond
a l'apparence d'une tache plus claire sur le fond sombre du cytoplasme
et est souvent situé au pôle cellulaire opposé
au support.
L'ostéoblaste est la cellule qui construit la lamelle
osseuse. Nous verrons plus loin que toute ostéogénèse
se fait contre un support. Le support de l'ostéogénèse
est représenté ici par une bande longitudinale colorée
en bleu. Les ostéoblastes se disposent contre ce support
en une couche continue de cellules cubiques. Leur cytoplasme est
intensément basophile, ce qui se traduit dans cette préparation
par une couleur violette. Cette basophilie est due à la
richesse en réticulum endoplasmique rugueux. Le noyau rond
a l'apparence d'une tache plus claire sur le fond sombre du cytoplasme
et est souvent situé au pôle cellulaire opposé
au support.
L'ostéoblaste a plusieurs fonctions. La principale est
de produire la phase organique de la lamelle osseuse qui, tant
qu'elle n'est pas calcifiée, s'appelle substance ostéoïde.
Il sécrète aussi des phosphatases alcalines. Ces
enzymes hydrolysent la gaine de polyphosphates qui enveloppe les
fibres collagènes et inhibe la calcification. L'ostéoblaste
intervient enfin dans la calcification elle-même. Il sécrète
les premiers éléments minéraux sous la forme
d'ions solubles qui précipitent en phase amorphe. Il intervient
enfin dans la régulation de l'ostéolyse; il peut
freiner l'activité des ostéoclastes en sécrétant
une prostaglandine; il peut aussi la renforcer en sécrétant
un facteur qui augmente la mobilité des ostéoclastes
et une collagénase qui détruit les fibres collagènes.
Les ostéoblastes proviennent de cellules mésenchymateuses
prédéterminées, appelées cellules
ostéoprogénitrices. Seule leur situation à
proximité d'autres cellules osseuses permet de les identifier
dans les préparations classiques. Elles se trouvent ici
dans le tissu conjonctif muqueux, entre le vaisseau sanguin et
la rangée d'ostéoblastes.
L'ostéoblaste est donc la cellule de l'os en formation.
Il élabore la phase organique, produit des phosphatases
alcalines et intervient dans la calcification et la régulation
de l'ostéolyse. Il provient d'une cellule mésenchymateuse.
Ostéocyte
 L'ostéoblaste, en sécrétant la phase organique,
s'enferme progressivement dans sa propre production. Il devient
alors un ostéocyte ou cellule de l'os formé. Chaque
ostéocyte est logé dans une petite cavité
ou lacune. Les lacunes à bords lisses contiennent des ostéocytes
jeunes qui achèvent le dépôt qu'ils avaient
entamé lorsqu'ils étaient encore ostéoblastes.
Les lacunes à bords plus irréguliers contiennent
des ostéocytes âgés.
L'ostéoblaste, en sécrétant la phase organique,
s'enferme progressivement dans sa propre production. Il devient
alors un ostéocyte ou cellule de l'os formé. Chaque
ostéocyte est logé dans une petite cavité
ou lacune. Les lacunes à bords lisses contiennent des ostéocytes
jeunes qui achèvent le dépôt qu'ils avaient
entamé lorsqu'ils étaient encore ostéoblastes.
Les lacunes à bords plus irréguliers contiennent
des ostéocytes âgés.
Les ostéocytes épousent la forme de leur cavité.
Leurs longs prolongements cytoplasmiques s'enfoncent dans les
canalicules qui, d'une lamelle à l'autre, unissent les
lacunes. Dans ces canalicules, les prolongements de cellules voisines
sont unis par des jonctions communicantes et enveloppés,
comme l'ostéocyte lui-même, d'un revêtement
externe, représenté en rouge dans le schéma.
La masse du tissu osseux est ainsi occupée par un réseau
cellulaire important.
L'ostéocyte a une double fonction. L'ostéocyte jeune
termine la synthèse entamée par l'ostéoblaste.
L'ostéocyte âgé exerce, au moyen de son système
lysosomial, une activité ostéolytique importante.
Cette double fonction est sous le contrôle hormonal de la
parathormone et de la calcitonine. Les ostéocytes interviennent
donc dans les échanges ioniques.
L'ostéocyte est donc la cellule de l'os formé.
Il est inclus dans une logette et ses nombreux prolongements s'insinuent
dans les canalicules de la lamelle osseuse. Il participe à
l'ostéogénèse et à l'ostéolyse.
Ostéoclaste
 Dans cette travée osseuse photographiée à
faible grossissement, les points clairs sont des lacunes ostéocytaires.
Le bord inférieur de la travée est limité
par une ligne bleue de substance ostéoïde contre laquelle
sont alignés des ostéoblastes. Le bord supérieur
est marqué par plusieurs érosions occupées
par de grandes cellules multinucléées. Ce sont des
ostéoclastes et les dépressions creusées
dans la travée sont des lacunes de Howship.
Dans cette travée osseuse photographiée à
faible grossissement, les points clairs sont des lacunes ostéocytaires.
Le bord inférieur de la travée est limité
par une ligne bleue de substance ostéoïde contre laquelle
sont alignés des ostéoblastes. Le bord supérieur
est marqué par plusieurs érosions occupées
par de grandes cellules multinucléées. Ce sont des
ostéoclastes et les dépressions creusées
dans la travée sont des lacunes de Howship.
L'ostéoclaste, vu à plus fort grossissement, coiffe
une lame osseuse et s'applique contre elle à la manière
d'une ventouse. La région de la cellule proche de l'os
est claire parce que remplie de vacuoles. Les noyaux sont tassés
du côté opposé.
Du côté de la travée osseuse, la membrane
plasmique de l'ostéoclaste a l'aspect d'une bordure en
brosse. Elle est formée de nombreux replis délimitant
les canalicules qui s'enfoncent à l'intérieur du
cytoplasme. L'ostéoclaste libère par exocytose des
hydrolases acides et les protons nécessaires à l'activité
de ces enzymes. Les protons dissolvent les cristaux d'apatite,
puis dissocient les fibres collagènes qui sont ensuite
détruites par les hydrolases. Les produits de dégradation
sont endocytés par la cellule et les ions sont remis en
circulation. L'ostéoclaste participe ainsi au maintien
du taux normal du calcium et des phosphates dans le plasma sanguin.
Cette fonction est contrôlée par la parathormone
et la calcitonine.
Les ostéoclastes sont donc des cellules géantes
plurinucléées, responsables de la résorption
osseuse massive. Ils ont leur origine dans la moelle osseuse et
proviennent d'une lignée particulière de monocytes.
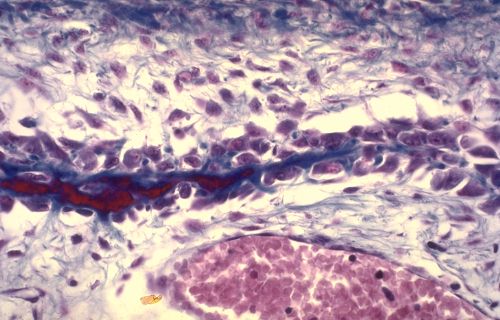
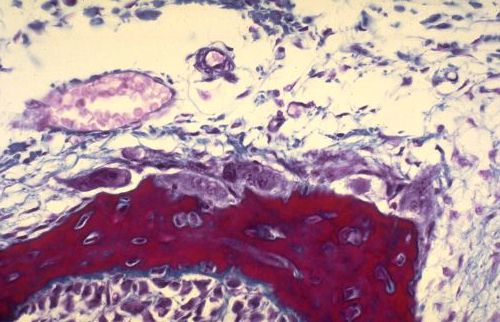
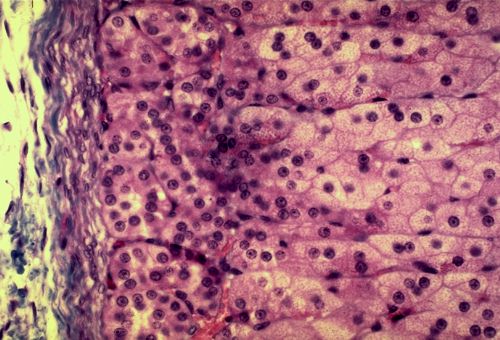
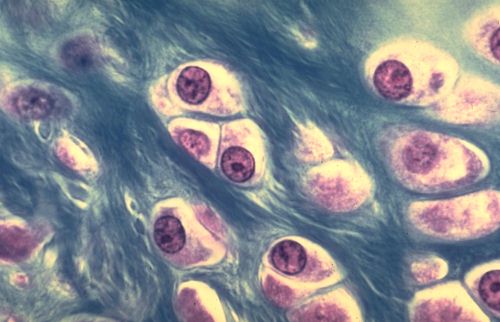
Vous terminerez votre étude en analysant les quatre dernières
images. De quel(s) tissu(s) s'agit-il et pourquoi?
 Réponse
Réponse
 Réponse
Réponse
 Réponse
Réponse
 Réponse
Réponse
Leçon suivante: Elaboration d'une lamelle osseuse
 Les deux composants du tissu
réticulé peuvent être
mis en évidence par l'éosine, qui colore les cellules
en rouge, et par une imprégnation à l'argent, qui
colore en noir les fibres collagènes de type III. Les fibres
dessinent un réseau à larges mailles ouvertes qui
contiennent de petites cellules sanguines rondes. Les cellules
réticulées sont accrochées aux fibres. Elles
sont allongées et leurs contours sont mal définis.
L'une d'entre elles est désignée par une flèche.
Les deux composants du tissu
réticulé peuvent être
mis en évidence par l'éosine, qui colore les cellules
en rouge, et par une imprégnation à l'argent, qui
colore en noir les fibres collagènes de type III. Les fibres
dessinent un réseau à larges mailles ouvertes qui
contiennent de petites cellules sanguines rondes. Les cellules
réticulées sont accrochées aux fibres. Elles
sont allongées et leurs contours sont mal définis.
L'une d'entre elles est désignée par une flèche.