2. Les tissus conjonctifs
2.1. Définition
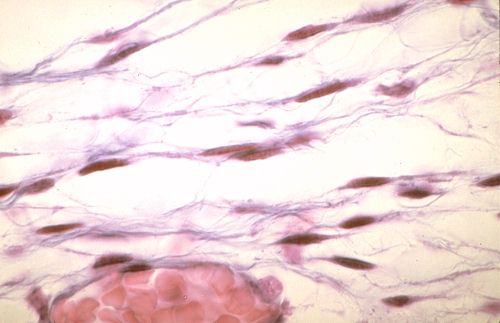
 Les tissus épithéliaux et les tissus conjonctifs
sont très différents. Dans les épithéliums,
les cellules sont étroitement associées sans interposition
de structure. Dans un tissu conjonctif, comme celui illustré
ici, les cellules, dont on ne voit que le noyau, sont séparées
par de nombreuses fibres, colorées en bleu-vert. Les espaces
entre les cellules et les fibres contiennent une substance qui,
dans des préparations comme celle-ci, n'est pas colorée.
Ce tissu est un tissu conjonctif.
Les tissus épithéliaux et les tissus conjonctifs
sont très différents. Dans les épithéliums,
les cellules sont étroitement associées sans interposition
de structure. Dans un tissu conjonctif, comme celui illustré
ici, les cellules, dont on ne voit que le noyau, sont séparées
par de nombreuses fibres, colorées en bleu-vert. Les espaces
entre les cellules et les fibres contiennent une substance qui,
dans des préparations comme celle-ci, n'est pas colorée.
Ce tissu est un tissu conjonctif.
Dans les tissus conjonctifs, les cellules sont éparpillées
et séparées les unes des autres par des fibres et
de la substance fondamentale dont l'ensemble forme la matrice
extracellulaire.
Tous les tissus conjonctifs répondent à cette définition,
ils n'ont pas tous la même structure. Parmi eux, on distingue
le tissu conjonctif non spécialisé ou très
peu spécialisé et les tissus conjonctifs particuliers,
très différenciés, que sont:
2.2. Tissus conjonctifs non spécialisés
2.2.1. Cellules
 Les cellules du tissu conjonctif non spécialisé
sont les cellules mésenchymateuses et les fibroblastes.
Les cellules du tissu conjonctif non spécialisé
sont les cellules mésenchymateuses et les fibroblastes.
Les cellules mésenchymateuses sont particulièrement
abondantes chez l'embryon. Elles sont avant tout des cellules
conjonctives peu différenciées, elles possèdent
de longs prolongements grâce auxquels elles forment la trame
du mésenchyme embryonnaire. Ces cellules mésenchymateuses
sont petites, allongées ou étoilées. Leur
noyau ovoïde est central; le cytoplasme est peu abondant.
Si les cellules mésenchymateuses ont toutes la même
morphologie chez l'embryon, elles n'ont pas toutes la même
destinée. Au cours du développement, les unes se
différencieront en fibroblastes, d'autres en cellules conjonctives
d'un autre type, d'autres encore en cellules sanguines. Ces différentes
populations sont "prédéterminées",
très tôt au cours de la vie embryonnaire. Elles se
différencient sous l'influence d'inducteurs spécifiques.
Des cellules mésenchymateuses persistent chez l'adulte.
Elles se nomment péricytes lorsqu'elles sont situées
près des capillaires (C). Dans leur noyau, la chromatine
est condensée en blocs. Leur cytoplasme est peu abondant
et contient quelques mitochondries, un peu de réticulum
endoplasmique rugueux et de nombreux polyribosomes. L'endothélium
des capillaires est séparé des péricytes
par la membrane basale, indiquée par des têtes de
flèches. Quelques prolongements d'autres péricytes,
indiqués par des flèches, s'insinuent entre cette
membrane basale et la cellule mésenchymateuse. Les péricytes
peuvent se différencier en fibroblastes ou en cellules
adipeuses.
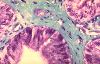 Les cellules les plus fréquentes dans un tissu conjonctif
adulte non spécialisé sont les fibroblastes. Dans
ce chorion, ils sont éparpillés entre les autres
éléments du tissu et, à ce grossissement,
seul leur noyau est visible. Les fibroblastes vus à plus
fort grossissement sont fusiformes. Leur noyau est allongé
dans l'axe de la cellule. Le cytoplasme est légèrement
basophile. Le degré de cette basophilie dépend de
la quantité de réticulum endoplasmique rugueux.
Les cellules les plus fréquentes dans un tissu conjonctif
adulte non spécialisé sont les fibroblastes. Dans
ce chorion, ils sont éparpillés entre les autres
éléments du tissu et, à ce grossissement,
seul leur noyau est visible. Les fibroblastes vus à plus
fort grossissement sont fusiformes. Leur noyau est allongé
dans l'axe de la cellule. Le cytoplasme est légèrement
basophile. Le degré de cette basophilie dépend de
la quantité de réticulum endoplasmique rugueux.
L'aspect caractéristique du fibroblaste en microscopie
électronique est son abondance en réticulum endoplasmique
rugueux (R). Dans son noyau, la chromatine est éparse et
le nucléole (N) a un aspect spongieux. Les structures en
dehors de la cellule sont des fibres collagènes (Co) que
nous étudierons en détail plus loin. Voici le schéma
de l'ultrastructure du fibroblaste. Il a tous les caractères,
représentés en noir dans le schéma, d'une
cellule productrice de protéines : l'important réticulum
endoplasmique rugueux (R), mais aussi un système de Golgi
très développé et de nombreuses vésicules
d'exocytose qui s'abouchent à la membrane plasmique. Mais
il possède aussi les caractères, représentés
en rouge, d'une cellule qui endocyte : de nombreuses vésicules
d'endocytose et un grand nombre de lysosomes (L).
Cette ultrastructure démontre la double fonction
des fibroblastes. Ils ont d'abord une activité
de synthèse et sécrètent les fibres et la
substance fondamentale du tissu conjonctif. Mais ils produisent
aussi les enzymes nécessaires à la dégradation
des protéines de la matrice extracellulaire. Les fibroblastes
endocytent les résidus de cette dégradation et,
lorsque cette activité prédomine, les caractères
liés à l'endocytose sont plus importants. L'ensemble
des fibroblastes d'un tissu conjonctif assure ainsi son renouvellement
constant.
2.2.2. Fibres
Le deuxième constituant des tissus conjonctifs est l'ensemble
des fibres, parmi lesquelles on distingue les fibres collagènes
et les fibres élastiques.
2.2.2.1. Structure des fibres de collagène
 Les fibres collagènes sont les plus abondantes.
Dans cette préparation de peau colorée selon une
méthode trichromique, elles sont bleues. Elles sont groupées
en faisceaux entre les cellules. Ces fibres sont des assemblages
de nombreuses fibrilles provenant elles-mêmes de l'association
de molécules, les protofibrilles de tropocollagène.
Les fibres collagènes sont les plus abondantes.
Dans cette préparation de peau colorée selon une
méthode trichromique, elles sont bleues. Elles sont groupées
en faisceaux entre les cellules. Ces fibres sont des assemblages
de nombreuses fibrilles provenant elles-mêmes de l'association
de molécules, les protofibrilles de tropocollagène.
La protofibrille de tropocollagène mesure 280 nm
en longueur et 1 à 2 nm en épaisseur. C'est un trimère,
composé de trois monomères ou chaînes alpha.
Chaque chaîne (A) est un polypeptide hélicoïdal,
dont chaque spire mesure environ un nm et contient 3 acides aminés.
Les trois monomères spiralés sont à leur
tour enroulés les uns autour des autres en formant une
hélice (B) dont chaque tour de spire mesure environ 8,6
nm et contient 10 acides aminés. Leurs extrémités
ne sont pas spiralées.
Dans la cellule, la production de collagène débute
par la synthèse d'un précurseur, le procollagène.
Les chaînes du procollagène sont produites individuellement
dans le réticulum endoplasmique rugueux et associées
en trimères hélicoïdaux dans le complexe de
Golgi. Ceux-ci sont transportés dans des vésicules
golgiennes (VG) vers la membrane plasmique et exocytés.
Pendant tout ce trajet intracellulaire, les trois monomères
conservent à leurs deux extrémités un court
segment non spiralé qui empêche la polymérisation
intracellulaire des trimères. L'excision de ces extrémités
dans le milieu extracellulaire transforme le procollagène
en tropocollagène et permet l'association des protofibrilles.
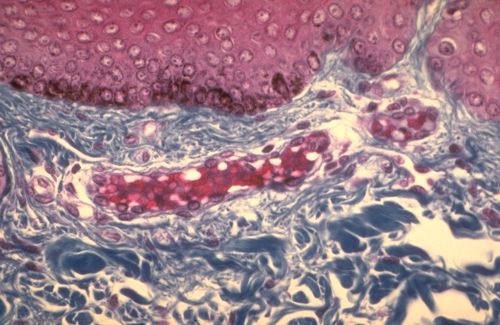
2.2.2.2. Principaux types de collagène
La séquence des acides aminés dans les chaînes
détermine le type de collagène. Le collagène
de type I est le plus répandu. Il entre dans la
constitution du derme cutané, de l'os, du cartilage fibreux
et du tendon. Il forme de gros faisceux fibrillaires comme
ceux que nous avons eu l'occasion de voir dans différentes
images. Le collagène de type I est coloré en jaune
, orange ou rouge lorsqu'il est, comme ici, coloré au rouge
Sirius et examiné en lumière polarisée.
Le collagène de type II est caractéristique
du cartilage hyalin. Le rouge Sirius le colore de façon
variable. Il ne forme pas de gros faisceaux mais un très
fin feutrage de petites fibres bien visibles dans une micrographie
électronique comme celle-ci. C'est parce qu'il ne forme
pas de faisceaux que la matrice cartilagineuse a un aspect homogène
lorsqu'elle est vue à faible grossissement.
Le collagène de type III correspond à ce
que l'on appelait la réticuline. Ses fibres se disposent
en réseau, d'où leur nom de fibres réticulées.
Il a une couleur verte lorsqu'il est coloré au rouge Sirius
et examiné en lumière polarisée. Il est observé
ici dans du tissu adipeux.
Les fibres réticulées ne sont pas visibles dans
les préparations habituelles. Pour les mettre en évidence,
on utilise des sels d'argent qui les colorent en noir. Les fibres
réticulées forment la trame des organes hématopoïétiques;
on les trouve aussi autour des cellules adipeuses et musculaires
lisses.
Le collagène de type IV n'existe que dans les membranes
basales. Il ne forme pas de fibres, probablement
parce que les extrémités non spiralées ne
sont pas excisées après la sécrétion.
L'exemple choisi est celui de la membrane basale du filtre rénal,
photographiée à très fort grossissement.
Elle est située entre l'endothélium qui limite la
lumière vasculaire dans le bas de l'image et l'épithélium
du glomérule. Les deux tissus sont particuliers; l'endothélium
est percé de pores et l'épithélium ménage
des fentes qui permettent la filtration.
En microscopie électronique, l'association des protofibrilles
de collagène de type I et III produit dans les fibres une
alternance de bandes claires et sombres qui leur donne un aspect
strié. La périodicité de cette striation
transversale est de 64 nm.
En effet, comme le montre le schéma d'une fibre, les molécules
de tropocollagène sont séparées par un espace
d'environ 30 nm et donc décalées les unes par rapport
aux autres. Ce décalage laisse libre une de leurs extrémités
et permet ainsi la formation de fibres d'épaisseur et de
longueur virtuellement indéterminées. De plus, les
fibres sont reliées les unes aux autres par des liaisons
covalentes latérales situées à leurs extrémites.
La superposition de ces espaces vides dans les fibrilles correspond
à des zones moins denses aux électrons. Les zones
sombres régulièrement espacées d'une distance
de 64 nm correspondent aux régions voisines. La très
forte résistance mécanique du collagène résulte
des liaisons latérales covalentes : un faisceau de fibres
d'un diamètre d'un millimètre peut résister
à une tension de près de 10 kg.
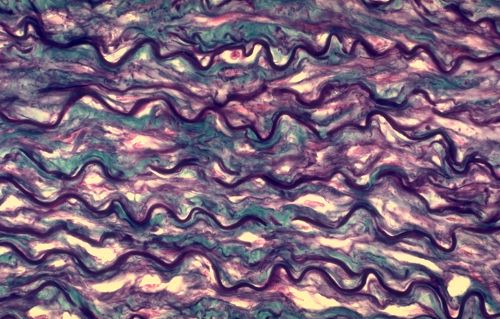
2.2.2.3. Fibres élastiques
 Les fibres élastiques sont moins fréquentes que
les fibres collagènes. Elles peuvent être étirées
et revenir spontanément à leur longueur initiale.
Elles sont abondantes dans les organes soumis à des variations
importantes de volume, tels que les grosses artères et
les poumons. On les trouve aussi dans certains tendons et ligaments,
dans le derme cutané et dans le cartilage élastique.
Cette paroi artérielle en contient beaucoup, elles sont
groupées en lames ondulées, elles sont colorées
en violet par l'aldéhyde-fuchsine. Les quelques faisceaux
de fibres collagènes, entre les lames élastiques,
sont colorés en bleu.
Les fibres élastiques sont moins fréquentes que
les fibres collagènes. Elles peuvent être étirées
et revenir spontanément à leur longueur initiale.
Elles sont abondantes dans les organes soumis à des variations
importantes de volume, tels que les grosses artères et
les poumons. On les trouve aussi dans certains tendons et ligaments,
dans le derme cutané et dans le cartilage élastique.
Cette paroi artérielle en contient beaucoup, elles sont
groupées en lames ondulées, elles sont colorées
en violet par l'aldéhyde-fuchsine. Les quelques faisceaux
de fibres collagènes, entre les lames élastiques,
sont colorés en bleu.
Les fibres élastiques sont produites par les fibroblastes
et les cellules musculaires lisses. Dans cette micrographie électronique
d'une artère, une lame élastique est située
sous l'endothélium et est désignée par une
flèche. Il en existe une autre, désignée
par une tête de flèche, dans la partie inférieure
de l'image. Les cellules entre les deux lames élastiques
sont des cellules musculaires lisses.
Les deux lames de l'image précédente paraissaient
homogènes. Une telle lame, photographiée à
plus fort grossissement, se trouve au centre de l'image. On peut
observer son microsquelette de très petites fibrilles soudées
par une substance amorphe, l'élastine. La lame élastique
est entourée de fibres collagènes, coupées
transversalement dans le haut de l'image et longitudinalement
en bas et à droite.
En résumé, les fibres collagènes de type
I sont généralement groupées en faisceaux.
Elles présentent une striation périodique de 64
nm. Les fibres collagènes de type III sont cependant associées
en réseau et s'appellent pour cette raison fibres réticulées;
elles sont mises en évidence par une coloration à
l'argent. Les fibres élastiques sont généralement
groupées en lames. Elles sont colorées spécifiquement
par l'aldéhyde fuchsine et l'orcéine.
2.2.3. Substance fondamentale
2.2.3.1. Composition
Cellules et fibres baignent dans la substance fondamentale, partie
amorphe de la matrice extracellulaire. La substance fondamentale
est un gel très hydraté dont la consistance
assure la turgescence des tissus conjonctifs. Elle est aussi responsable
de la cohésion des éléments figurés,
cellules et fibres. Elle est enfin, à cause de son contenu
aqueux, le lieu des échanges entre les cellules et le sang.
Ces différentes propriétés de la substance
fondamentale s'expliquent par leur contenu en protéoglycanes
et en glycoprotéines très particulières,
les fibronectines.
Les protéoglycanes sont des molécules dont la longueur
peut atteindre plusieurs microns. Chaque protéoglycane
est composé d'un axe protéique auquel se lient de
façon covalente de très nombreux glycosaminoglycanes.
Chaque glycosaminoglycane est une longue chaîne glucidique
dont l'unité de base est un disaccharide comprenant un
groupement aminé et un groupement uronique souvent substitués.
Ces chaînes sont relativement rigides de sorte que la totalité
de la molécule occupe une place considérable. Elles
sont aussi très hydrophiles; chaque protéoglycane
retient donc dans ses mailles une quantité d'eau importante.
Ces deux particularités expliquent la consistance gélatineuse
et la turgescence de la substance fondamentale; elles expliquent
aussi pourquoi les ions et les métabolites de faible poids
moléculaire diffusent très aisément dans
les tissus conjonctifs.
La turgescence de la substance fondamentale est encore amplifiée
parce que les protéoglycanes forment des agrégats.
Ils se lient pour cela à l'acide hyaluronique. Cet
acide est est une très longue chaîne composée
de plusieurs milliers de résidus osidiques. Cette chaîne
peut fixer, de façon non covalente et par l'intermédiaire
de petites protéines, plusieurs centaines de protéoglycanes.
Les agrégats ainsi constitués sont gigantesques,
ont un poids moléculaire de plus de 100 millions et la
taille d'une bactérie.
La cohésion entre les éléments figurés,
cellules et fibres, est assurée par les fibronectines.
Ce sont des molécules composées de deux chaînes
reliées à l'une de leurs extrémités
par des ponts disulfures. Elles se lient, d'une part, au collagène
et aux protéoglycanes. D'autre part, elles se fixent à
la membrane des cellules et sont en rapport, par l'intermédiaire
de protéines membranaires, avec les faisceaux de microfilaments
intracellulaires d'actine.
2.2.3.2. Colorations
 On peut, en microscopie optique, apprécier la richesse
de la substance fondamentale en chaînes glucidiques par
plusieurs méthodes. Cette préparation a été
traitée par la technique P.A.S., initiales qui signifient
Periodic Acid Schiff. La substance fondamentale de tous les tissus
conjonctifs, en particulier celle des trois gros nodules cartilagineux
au centre de l'image, est colorée en rouge violet. Le mucus
des cellules caliciformes incluses dans l'épithélium
pseudostratifiéqui borde la cavité située
dans le haut de l'image, l'est aussi.
On peut, en microscopie optique, apprécier la richesse
de la substance fondamentale en chaînes glucidiques par
plusieurs méthodes. Cette préparation a été
traitée par la technique P.A.S., initiales qui signifient
Periodic Acid Schiff. La substance fondamentale de tous les tissus
conjonctifs, en particulier celle des trois gros nodules cartilagineux
au centre de l'image, est colorée en rouge violet. Le mucus
des cellules caliciformes incluses dans l'épithélium
pseudostratifiéqui borde la cavité située
dans le haut de l'image, l'est aussi.
Voici le principe de la technique du PAS. L'acide periodique est
un oxydant puissant qui rompt les cycles osidiques, produisant
pour chaque cycle deux groupes aldéhydes à partir
des fonctions hydroxyles, si celles-ci ne sont pas substituées.
Les aldéhydes sont ensuite combinés au réactif
de Schiff, fuchsine décolorée au dioxyde de soufre.
Cette combinaison produit une substance rouge. On peut ainsi détecter
de longues chaînes osidiques ou des protéines glycosylées,
mais on ne peut détecter les oses simples qui sont solubles
et donc extraits par la préparation.
On peut aussi mettre les glycosaminoglycanes en évidence
grâce à la densité de leurs radicaux acides.
Cette densité est mise en évidence par un colorant
métachromatique comme le bleu de toluidine. Ce colorant
bleu colore la substance fondamentale de ce tissu conjonctif cartilagineux
en violet, parce que la densité des radicaux acides y est
importante. Les autres structures qui n'ont pas cette particularité
sont colorées en bleu. L'obtention d'une couleur violette
avec un colorant basique bleu porte le nom de métachromasie.
On peut expliquer schématiquement le
principe de la métachromasie
de la façon suivante. Les colorants métachromatiques,
comme tous les colorants basiques, se fixent aux groupements acides
des glycosaminoglycanes. Lorsque la densité de ces groupements
est faible (A), les molécules du colorant restent isolées
les unes des autres et les structures sont colorées en
bleu. Lorsque la densité de ces groupements acides est
élevée (B), les molécules de colorant qui
s'y fixent sont tellement proches qu'elles se recombinent entre
elles, ce qui modifie leur spectre d'absorption et explique le
passage de la couleur bleue à la couleur rouge violet.
2.2.4. Types de tissus conjonctifs
D'après la proportion de ces constituants, on distingue:
2.2.4.1. Muqueux
 Dans ce tissu conjonctif muqueux,
les cellules mésenchymateuses
sont colorées en rose, et les fibres en bleu. Les nombreux
espaces blancs entre les cellules contiennent de la substance
fondamentale qui n'est pas mise en évidence. Les cellules
sont souvent en contact les unes avec les autres par leurs prolongements.
Les fibres forment de minces faisceaux. Le tissu conjonctif
muqueux est un tissu conjonctif, où prédomine la
substance fondamentale. Très important chez l'embryon,
il se différenciera plus tard en d'autres variétés
de tissus. Chez l'adulte, le tissu conjonctif muqueux est rare;
il existe autour de quelques glandes et de quelques vaisseaux.
Dans ce tissu conjonctif muqueux,
les cellules mésenchymateuses
sont colorées en rose, et les fibres en bleu. Les nombreux
espaces blancs entre les cellules contiennent de la substance
fondamentale qui n'est pas mise en évidence. Les cellules
sont souvent en contact les unes avec les autres par leurs prolongements.
Les fibres forment de minces faisceaux. Le tissu conjonctif
muqueux est un tissu conjonctif, où prédomine la
substance fondamentale. Très important chez l'embryon,
il se différenciera plus tard en d'autres variétés
de tissus. Chez l'adulte, le tissu conjonctif muqueux est rare;
il existe autour de quelques glandes et de quelques vaisseaux.
2.2.4.2. Lâche
 Ce deuxième tissu conjonctif est le chorion du feuillet
glandulaire de l'estomac. Ses fibres, colorées en bleu,
forment de fins faisceaux entrelacés. Les cellules sont
nombreuses; leur noyau est ovale; le cytoplasme est rarement visible
de part et d'autre du noyau. Certaines sont des fibroblastes,
d'autres sont des cellules étrangères qui ont envahi
le tissu conjonctif. La substance fondamentale est relativement
peu importante. Ce tissu conjonctif, où la composante
cellulaire est particulièrement importante, est un tissu
conjonctif lâche.
Ce deuxième tissu conjonctif est le chorion du feuillet
glandulaire de l'estomac. Ses fibres, colorées en bleu,
forment de fins faisceaux entrelacés. Les cellules sont
nombreuses; leur noyau est ovale; le cytoplasme est rarement visible
de part et d'autre du noyau. Certaines sont des fibroblastes,
d'autres sont des cellules étrangères qui ont envahi
le tissu conjonctif. La substance fondamentale est relativement
peu importante. Ce tissu conjonctif, où la composante
cellulaire est particulièrement importante, est un tissu
conjonctif lâche.
2.2.4.3. Dense
 Dans le tissu conjonctif dense, les fibres sont particulièrement
nombreuses, la substance fondamentale est relativement peu
abondante et quelques fibroblastes sont perdus dans la masse des
faisceaux fibrillaires. D'après l'agencement de ces faisceaux,
le tissu conjonctif dense est régulier ou irrégulier.
Dans les tissus conjonctifs denses réguliers, les faisceaux
et les fibres collagènes ont une orientation bien définie.
La disposition de ces faisceaux est cependant différente
dans un tendon ou un ligament et dans la cornée.
Dans le tissu conjonctif dense, les fibres sont particulièrement
nombreuses, la substance fondamentale est relativement peu
abondante et quelques fibroblastes sont perdus dans la masse des
faisceaux fibrillaires. D'après l'agencement de ces faisceaux,
le tissu conjonctif dense est régulier ou irrégulier.
Dans les tissus conjonctifs denses réguliers, les faisceaux
et les fibres collagènes ont une orientation bien définie.
La disposition de ces faisceaux est cependant différente
dans un tendon ou un ligament et dans la cornée.
Voici la coupe longitudinale d'un tendon, qui relie un muscle
à une pièce osseuse. Un ligament, qui relie deux
os, aurait la même structure. Le tissu conjonctif dense
est très régulier car il est composé d'épais
faisceaux de fibres collagènes, parallèles et colorés
en bleu. Dans ces faisceaux, les fibroblastes sont disposés
parallèlement aux fibres. Les différents faisceaux
de collagène sont unis par du tissu conjonctif lâche,
plus clair, très cellulaire et pauvre en collagène.
La grande résistance des tendons et des ligaments provient
de l'orientation des fibres et des faisceaux dans le sens de la
traction.
Dans la cornée, le chorion de l'épithélium
pavimenteux stratifié épidermoïde est un tissu
conjonctif dense régulier dont les fibres collagènes
sont disposées en couches superposées. La substance
fondamentale est localisée dans des espaces entre les couches.
Les cellules sont appliquées contre les fibres, à
la limite de ces interstices. Dans chaque couche, les fibres sont
toutes parallèles mais celle d'une couche sont disposées
perpendiculairement à celles des couches voisines. Elles
ont la forme de stries dans les couches où elles sont coupées
longitudinalement et celle de points dans les couches où
elles sont coupées transversalement.
Le tissu conjonctif dense est donc un tissu conjonctif où
les fibres prédominent. Le plus commun est irrégulier,
les faisceaux de fibres étant disposés sans ordre
apparent. Dans les tendons et les ligaments, il est régulier
et les fibres collagènes sont disposées en faisceaux
parallèles. Dans la cornée et les aponévroses,
il est régulier et les fibres collagènes sont agencées
en lames.
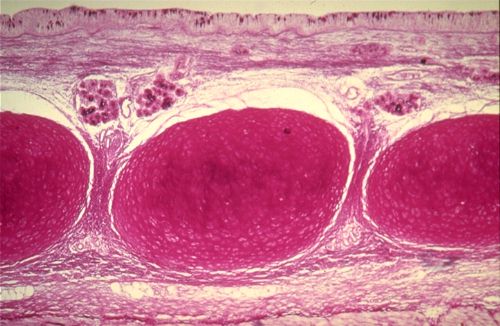
2.2.4.4. Elastique
 Dans le tissu conjonctif élastique, les fibres prédominent.
La plupart cependant sont disposées en lames parallèles,
colorées dans cette artère en rose. Les espaces
entre ces lames contiennent de la substance fondamentale et des
noyaux qui appartiennent à des cellules musculaires lisses.
Cette coloration ne permet cependant pas d'identifier la nature
des lames conjonctives. Les fibres élastiques, colorées
en violet par l'aldehyde fuchsine , sont associées en lames
épaisses, sinueuses, parallèles à la lumière
et anastomosées entre elles par des lames plus grêles.
Sa richesse en fibres élastiques fait de ce tissu conjonctif
un véritable tissu élastique. Chez l'homme, on le
trouve dans le ligament jaune de la colonne vertébrale,
dans les cordes vocales et dans des artères de gros calibre,
appelées pour cette raison artères élastiques.
Dans le tissu conjonctif élastique, les fibres prédominent.
La plupart cependant sont disposées en lames parallèles,
colorées dans cette artère en rose. Les espaces
entre ces lames contiennent de la substance fondamentale et des
noyaux qui appartiennent à des cellules musculaires lisses.
Cette coloration ne permet cependant pas d'identifier la nature
des lames conjonctives. Les fibres élastiques, colorées
en violet par l'aldehyde fuchsine , sont associées en lames
épaisses, sinueuses, parallèles à la lumière
et anastomosées entre elles par des lames plus grêles.
Sa richesse en fibres élastiques fait de ce tissu conjonctif
un véritable tissu élastique. Chez l'homme, on le
trouve dans le ligament jaune de la colonne vertébrale,
dans les cordes vocales et dans des artères de gros calibre,
appelées pour cette raison artères élastiques.
Leçon suivante: 2.3. Les tissus conjonctifs spécialisés
: tissu réticulé
 Les tissus épithéliaux et les tissus conjonctifs
sont très différents. Dans les épithéliums,
les cellules sont étroitement associées sans interposition
de structure. Dans un tissu conjonctif, comme celui illustré
ici, les cellules, dont on ne voit que le noyau, sont séparées
par de nombreuses fibres, colorées en bleu-vert. Les espaces
entre les cellules et les fibres contiennent une substance qui,
dans des préparations comme celle-ci, n'est pas colorée.
Ce tissu est un tissu conjonctif.
Les tissus épithéliaux et les tissus conjonctifs
sont très différents. Dans les épithéliums,
les cellules sont étroitement associées sans interposition
de structure. Dans un tissu conjonctif, comme celui illustré
ici, les cellules, dont on ne voit que le noyau, sont séparées
par de nombreuses fibres, colorées en bleu-vert. Les espaces
entre les cellules et les fibres contiennent une substance qui,
dans des préparations comme celle-ci, n'est pas colorée.
Ce tissu est un tissu conjonctif.