3. Le sang et les systèmes de défense (troisième
partie)
3.5. Hématopoïèse
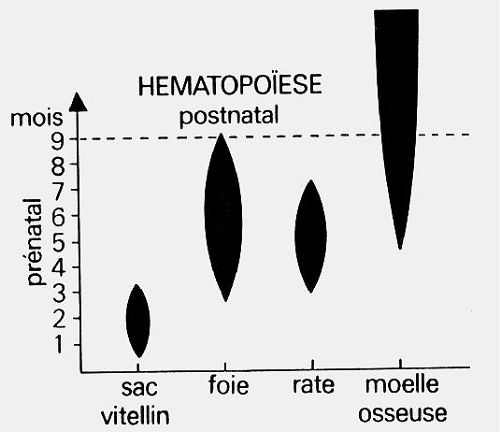
 L'hématopoïèse est continue tout au long de
la vie. Elle se fait dans les organes hématopoïétiques,
à partir de la différenciation de cellules mésenchymateuses.
Les organes hématopoïétiques changent en fonction
des périodes de la vie. Pendant la
période prénatale,
vers la deuxième semaine, l'hématopoïèse
débute dans le sac vitellin et produit uniquement des globules
rouges nucléés. Elle se poursuit dans le foie qui
produit des globules rouges anucléés ou érythrocytes,
des précurseurs des plaquettes et quelques leucocytes.
La rate est aussi le site de l'hématopoïèse,
pendant une période relativement brève. A partir
du quatrième mois, la moelle osseuse est l'organe hématopoïétique
principal; elle le demeure après la naissance.
L'hématopoïèse est continue tout au long de
la vie. Elle se fait dans les organes hématopoïétiques,
à partir de la différenciation de cellules mésenchymateuses.
Les organes hématopoïétiques changent en fonction
des périodes de la vie. Pendant la
période prénatale,
vers la deuxième semaine, l'hématopoïèse
débute dans le sac vitellin et produit uniquement des globules
rouges nucléés. Elle se poursuit dans le foie qui
produit des globules rouges anucléés ou érythrocytes,
des précurseurs des plaquettes et quelques leucocytes.
La rate est aussi le site de l'hématopoïèse,
pendant une période relativement brève. A partir
du quatrième mois, la moelle osseuse est l'organe hématopoïétique
principal; elle le demeure après la naissance.
La moelle osseuse est abondante dans les cavités de l'os
spongieux des os courts et des os plats, et celles des épiphyses
et des métaphyses des os longs. Elle est rare dans la cavité
diaphysaire des os longs. Dans cette métaphyse, les travées
d'os spongieux sont aisément reconnaissables. Elles sont
plus denses et plus rapprochées dans la partie droite;
la moelle par contre est plus abondante dans la partie gauche.
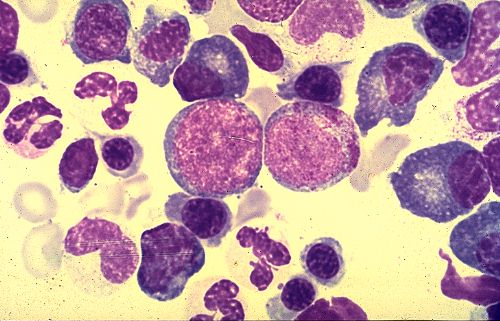
Dans cette coupe de moelle osseuse, examinée à faible
grossissement, Il est difficile d'identifier les différents
types cellulaires. On reconnaît néanmoins les précurseurs
des plaquettes sanguines, les mégacaryocytes, qui sont
volumineux et possèdent un noyau plurilobé. Les
plages claires représentent des adipocytes.
A plus fort grossissement, on peut reconnaître plusieurs
cellules sanguines différenciées et notamment des
polynucléaires et des lymphocytes. Parmi elles se trouvent
des cellules non différenciées. Celles qui sont
le plus aisément identifiables appartiennent à la
lignée des polynucléaires éosinophiles parce
que leur cytoplasme est rempli de granulations rouges. Le centre
de l'image est occupé par un large vaisseau dont l'endothélium
est très aplati et discontinu; c'est un sinusoïde
veineux, caractéristique de la moelle osseuse. A droite
de ce sinusoïde se trouve une grande cellule où l'on
reconnaît un noyau à plusieurs lobes; ici encore,
il s'agit d'un précurseur des plaquettes sanguines.
Les cellules mésenchymateuses, qui sont à l'origine
de la moelle, se différencient en cellules stromales et
en précurseurs des cellules sanguines. Les cellules stromales
comportent les cellules réticulées, les cellules
endothéliales et les adipocytes.
Dans la moelle osseuse, les cellules mésenchymateuses destinées
à se différencier en cellules sanguines sont appelées
cellules souches. Lorsqu'elles sont capables de donner naissance
à plusieurs types cellulaires, elles sont dites pluripotentes.
Les cellules souches pluripotentes se divisent et produisent des
cellules unipotentes, programmées pour produire chacune
un type cellulaire déterminé. Ces différents
types cellulaires peuvent être classés en différentes
lignées. La lignée lymphoïde produit les lymphocytes,
la lignée érythrocytaire produit les érythrocytes,
la lignée plaquettaire produit les plaquettes sanguines,
la lignée granulo-monocytaire produit les granulocytes
et les monocytes.
3.5.1. Lignée érythrocytaire.
 La cellule unipotente morphologiquement identifiable dans la lignée
érythrocytaire est le proérythroblaste. C'est une
volumineuse cellule. Son diamètre varie entre 14 et 20
µm. Le noyau est rond et central; sa chromatine est bien
dessinée. Le cytoplasme, riche en ribosomes, est très
basophile.
La cellule unipotente morphologiquement identifiable dans la lignée
érythrocytaire est le proérythroblaste. C'est une
volumineuse cellule. Son diamètre varie entre 14 et 20
µm. Le noyau est rond et central; sa chromatine est bien
dessinée. Le cytoplasme, riche en ribosomes, est très
basophile.
Le proérythroblaste se divise pour produire des érythroblastes
basophiles. Voici deux érythroblastes basophiles. Ils sont
plus petits que le proérythroblaste. Leur diamètre
varie entre 10 et 18 µm. La chromatine commence à
se condenser; les nucléoles sont visibles. Le cytoplasme
est basophile.
L'érythroblaste basophile se divise et, dans son cytoplasme,
apparaissent les premières traces d'un pigment, l'hémoglobine.
Il devient alors un érythroblaste polychromatophile, parce
que la coloration de son cytoplasme est intermédiaire entre
celle du basophile et celle de l'acidophile. L'érythroblaste
polychromatophile est plus petit; il se divise plusieurs fois.
Au fur et à mesure de ces divisions, sa chromatine se condense
et les nucléoles deviennent moins visibles; la basophilie
du cytoplasme diminue car la cellule ne synthétise plus
de ribosomes et la quantité d'hémoglobine augmente
progressivement.
Lorsque l'hémoglobine remplit tout le cytoplasme, l'érythroblaste
polychromatophile devient acidophile. Son diamètre varie
entre 7 et 14 µm. Son noyau est d'abord central et sa chromatine
est condensée en quelques blocs. Après quelques
divisions, le noyau devient pycnotique et excentrique. Le cytoplasme,
rempli d'hémoglobine, contient encore quelques ribosomes
et quelques mitochondries.
A la fin de cette évolution, le noyau est expulsé,
enveloppé d'un fin film de cytoplasme; il se trouve en
haut et à droite dans l'image. La portion anucléée
est alors un jeune globule rouge ou réticulocyte. Il sera
libéré dans la circulation sanguine.
Les érythrocytes proviennent donc de précurseurs
médullaires. Leur évolution, stimulée par
l'érythropoïétine, comporte plusieurs aspects.
- La taille de la cellule diminue.
- La basophilie du cytoplasme diminue
par disparition des ribosomes.
- L'acidophilie du cytoplasme augmente
par accumulation d'hémoglobine.
- Le noyau se condense et est finalement
expulsé.
Chez l'adulte normal, les réticulocytes représentent
moins de 1% du nombre total de globules rouges. Le contrôle
de la prolifération et de la différenciation des
précurseurs des érythrocytes est hormonal. Une baisse
de la concentration sanguine en oxygène ou une déperdition
sanguine stimule la sécrétion de l'érythropoïétine
par le rein. Celle-ci accélère la division et la
transformation des érythroblastes.
3.5.2. Lignée plaquettaire.
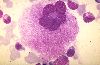 Le précurseur immédiat des plaquettes sanguines
est une très grande cellule, le mégacaryocyte. Son
noyau polyploïde est le résultat d'une suite d'endomitoses;
il est composé de nombreux lobes reliés entre eux
et disposés en couronne. Il provient d'une cellule souche
beaucoup plus petite et diploïde où la synthèse
et la duplication d'ADN n'ont pas été suivies de
caryocinèse, ni de cytocinèse, ce qui explique la
polyploïdie et le volume très important de la cellule,
que l'on apprécie particulièrement dans un frottis
médullaire. Ici, le cytoplasme est granuleux parce que
la cellule est presque au terme de son évolution.
Le précurseur immédiat des plaquettes sanguines
est une très grande cellule, le mégacaryocyte. Son
noyau polyploïde est le résultat d'une suite d'endomitoses;
il est composé de nombreux lobes reliés entre eux
et disposés en couronne. Il provient d'une cellule souche
beaucoup plus petite et diploïde où la synthèse
et la duplication d'ADN n'ont pas été suivies de
caryocinèse, ni de cytocinèse, ce qui explique la
polyploïdie et le volume très important de la cellule,
que l'on apprécie particulièrement dans un frottis
médullaire. Ici, le cytoplasme est granuleux parce que
la cellule est presque au terme de son évolution.
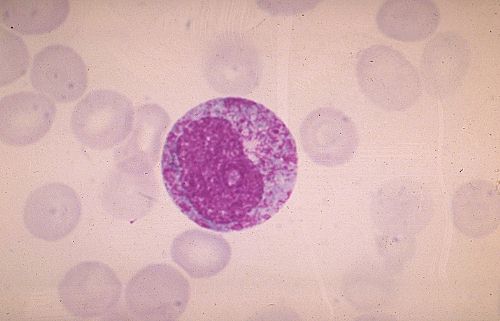
Dans une coupe de moelle osseuse, on reconnaît facilement
un mégacaryocyte même si l'on n'observe pas la totalité
de son volume. La cellule est grande; son cytoplasme est acidophile;
son noyau est plurilobé et beaucoup moins dense que le
noyau des petites cellules avoisinantes.
Le cytoplasme du mégacaryocyte est caractéristique.
Il est rempli de granules et possède un système
membranaire très développé. Les grains sont
ceux décrits dans les plaquettes sanguines; les plus nombreux
sont les grains alpha qui possèdent un nucléoïde;
les corps denses sont rares. Les nombreux saccules membranaires,
actuellement dispersés et sans orientation précise,
seront à l'origine d'un véritable système
de démarcation.
Lorsque le système de démarcation s'organise, le
mégacaryocyte est dit thrombocytogène, parce qu'il
est sur le point de libérer ses plaquettes. Les tubules
s'alignent et délimitent des "champs plaquettaires"
dont le centre est occupé par les granules.
Pour former les plaquettes sanguines, les mégacaryocytes
émettent des pseudopodes qui s'insinuent dans les interstices
des sinusoïdes veineux. Les membranes de démarcation
fusionnent avec la membrane plasmique et fragmentent ainsi le
cytoplasme. Ces fragments cytoplasmiques, libérés
dans la lumière vasculaire, sont les plaquettes sanguines.
La portion nucléée reste dans la moelle osseuse
et devient la proie des macrophages. Cette thrombocytopoïèse
est stimulée lorsque le nombre des plaquettes diminue dans
le sang.
L'origine des plaquettes sanguines est donc le mégacaryocyte
de la moelle osseuse, cellule très volumineuse au noyau
polyploïde. Les plaquettes sanguines proviennent de la fragmentation
de son cytoplasme, grâce à un système tubulaire
de démarcation.
3.5.3. Lignée granulo-monocytaire.
La lignée granulo-monocytaire produit les polynucléaires
neutrophiles, éosinophiles et basophiles et les monocytes.
Dans la lignée monocytaire, la cellule unipotente identifiable
dans un frottis médullaire est le monoblaste qui ressemble
au myéloblaste. Il se divise pour former des promonocytes.
Le promonocyte engendre des monocytes, qui ne se divisent plus,
passent dans le sang, puis dans les tissus où ils deviennent
des macrophages.
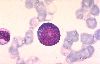 Dans la lignée granulocytaire, la cellule unipotente identifiable
dans un frottis médullaire est le myéloblaste. Son
diamètre varie entre 15 et 18 µm. Son noyau est excentrique,
accolé à la membrane plasmique, et les zones nucléolaires
y sont nettement délimitées. Le rapport nucléo-cytoplasmique
est élevé. Son cytoplasme est très basophile.
Dans la lignée granulocytaire, la cellule unipotente identifiable
dans un frottis médullaire est le myéloblaste. Son
diamètre varie entre 15 et 18 µm. Son noyau est excentrique,
accolé à la membrane plasmique, et les zones nucléolaires
y sont nettement délimitées. Le rapport nucléo-cytoplasmique
est élevé. Son cytoplasme est très basophile.
Le myéloblaste se divise pour donner des promyélocytes.
Le promyélocyte est une cellule plus grande. Son diamètre
varie entre 16 et 20 µm. Sa chromatine est bien dessinée.
Le rapport nucléo-cytoplasmique est plus faible qu'au stade
précédent. Son cytoplasme basophile contient des
grains azurophiles, de couleur rougeâtre. En évoluant,
le promyélocyte diminue de volume et son noyau se condense.
Le promyélocyte se divise pour donner des myélocytes.
Si le stade de promyélocyte est caractérisé
par la présence de grains azurophiles, celui de myélocyte
est caractérisé par la présence des grains
spécifiques neutrophiles, éosinophiles ou basophiles.
Le myélocyte basophile est évidemment rarissime.
Dans le myélocyte, le noyau est encore arrondi; il devient
un métamyélocyte lorsque son noyau est échancré.
Le myélocyte neutrophile est plus petit que le promyélocyte.
Son noyau est dense; le rapport nucléo-cytoplasmique est
faible. Le cytoplasme est légèrement basophile.
Il contient de nombreux grains spécifiques, à la
limite de la visibilité optique. Les grains plus épais
sont des grains azurophiles. Le myélocyte neutrophile devient
un métamyélocyte neutrophile.
Voici deux métamyélocytes neutrophiles. Ils sont
plus petits que le myélocyte. Le rapport nucléo-cytoplasmique
est faible; le noyau est échancré et très
condensé; les grains spécifiques neutrophiles donnent
au cytoplasme un aspect poussiéreux. La lobulation du noyau
se poursuivant, le métamyélocyte devient un polynucléaire
neutrophile qui passe dans la circulation sanguine.
Les myélocytes éosinophiles sont moins nombreux
que les myélocytes neutrophiles. Leur cytoplasme est masqué
par les nombreux grains spécifiques éosinophiles
et très volumineux.
Le métamyélocyte éosinophile est plus petit
que le myélocyte éosinophile. Son noyau est échancré,
sa chromatine très condensée. Son cytoplasme est
à peine visible car rempli de gros grains éosinophiles.
La lobulation du noyau se poursuivant, le métamyélocyte
devient un polynucléaire éosinophile qui passe dans
la circulation sanguine.
Tous les polynucléaires proviennent donc d'un précurseur
commun, le myéloblaste. Celui-ci, en se divisant, produit
des promyélocytes, dont le cytoplasme se garnit de grains
azurophiles. A leur tour, les promyélocytes produisent
des myélocytes caractérisés par la présence
de grains spécifiques. Les myélocytes, en se divisant,
produisent des métamyélocytes, au noyau échancré.
Ceux-ci ne se divisent plus et deviennent des polynucléaires,
au noyau plurilobé. Au cours de cette évolution,
les cellules deviennent de plus en plus petites, et la quantité
de réticulum endoplasmique rugueux et donc leur basophilie
diminuent.
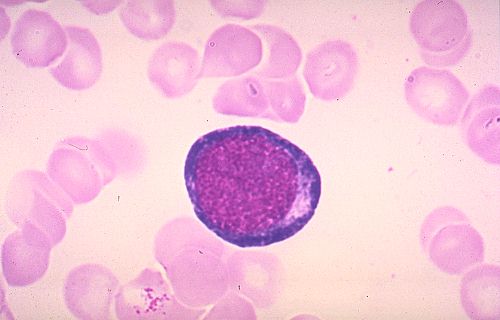
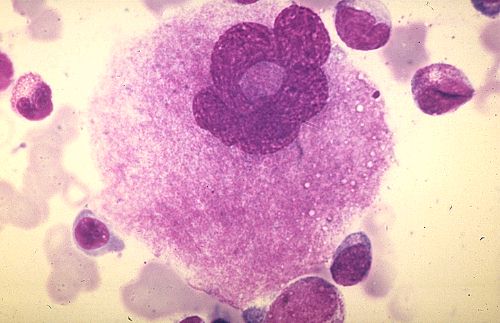
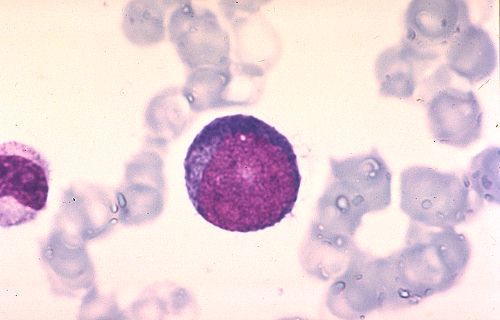
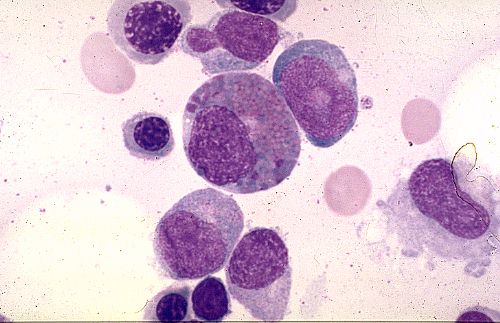
Pour terminer cette étude de la moelle osseuse et de l'hématopoièse,
essayez d'identifier les cellules représentées dans
les images suivantes.
 Réponse
Réponse
 Réponse
Réponse
 Réponse
Réponse
Les prochaines leçons seront consacrées au tissu
nerveux.
 L'hématopoïèse est continue tout au long de
la vie. Elle se fait dans les organes hématopoïétiques,
à partir de la différenciation de cellules mésenchymateuses.
Les organes hématopoïétiques changent en fonction
des périodes de la vie. Pendant la
période prénatale,
vers la deuxième semaine, l'hématopoïèse
débute dans le sac vitellin et produit uniquement des globules
rouges nucléés. Elle se poursuit dans le foie qui
produit des globules rouges anucléés ou érythrocytes,
des précurseurs des plaquettes et quelques leucocytes.
La rate est aussi le site de l'hématopoïèse,
pendant une période relativement brève. A partir
du quatrième mois, la moelle osseuse est l'organe hématopoïétique
principal; elle le demeure après la naissance.
L'hématopoïèse est continue tout au long de
la vie. Elle se fait dans les organes hématopoïétiques,
à partir de la différenciation de cellules mésenchymateuses.
Les organes hématopoïétiques changent en fonction
des périodes de la vie. Pendant la
période prénatale,
vers la deuxième semaine, l'hématopoïèse
débute dans le sac vitellin et produit uniquement des globules
rouges nucléés. Elle se poursuit dans le foie qui
produit des globules rouges anucléés ou érythrocytes,
des précurseurs des plaquettes et quelques leucocytes.
La rate est aussi le site de l'hématopoïèse,
pendant une période relativement brève. A partir
du quatrième mois, la moelle osseuse est l'organe hématopoïétique
principal; elle le demeure après la naissance.