Le tissu nerveux (Première partie)
4.1. Définition
Le système nerveux est l'ensemble des structures qui
coordonnent les fonctions d'un individu et permettent ses relations
avec le milieu extérieur.
Il est composé de cellules nerveuses spécialisées,
les neurones, et de cellules de soutien, les cellules gliales.
L'association des neurones et des cellules gliales est différente
dans le système nerveux périphérique et dans
le système nerveux central. Les cellules gliales du système
nerveux périphérique sont les cellules capsulaires
et les cellules de Schwann; celles du système nerveux central
sont les cellules épendymaires, les astrocytes et les oligodendrocytes.
Nous décrirons chaque type cellulaire en étudiant
la structure dans laquelle il se trouve.
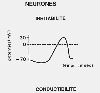 Les fonctions du système nerveux sont basées sur
deux propriétés cellulaires, l'irritabilité
et la conductibilité, particulièrement
développées dans le neurone. Son irritabilité
est sa capacité de réagir aux variations du milieu
extracellulaire en modifiant les propriétés électriques
de sa membrane. Lorsqu'aucun influx nerveux n'est transmis, le
potentiel électrique à l'intérieur de la
cellule est négatif d'environ -70 millivolts par rapport
à celui de l'extérieur; cette différence
de potentiel, ou potentiel de repos, est liée à
la différence de concentration en ions entre les deux milieux.
Lorsqu'un influx nerveux se déplace le long de la membrane
plasmique, la différence de potentiel transmembranaire
varie brusquement; l'intérieur de la cellule devient positif
(+30 millivolts) pendant quelques millisecondes puis reprend sa
valeur de repos de -70 millivolts. Cette brusque inversion est
appelée potentiel d'action. C'est la manifestation électrique
de l'influx nerveux. La conductibilité d'un neurone est
sa capacité de transmettre l'influx nerveux très
rapidement et sur de longues distances .
Les fonctions du système nerveux sont basées sur
deux propriétés cellulaires, l'irritabilité
et la conductibilité, particulièrement
développées dans le neurone. Son irritabilité
est sa capacité de réagir aux variations du milieu
extracellulaire en modifiant les propriétés électriques
de sa membrane. Lorsqu'aucun influx nerveux n'est transmis, le
potentiel électrique à l'intérieur de la
cellule est négatif d'environ -70 millivolts par rapport
à celui de l'extérieur; cette différence
de potentiel, ou potentiel de repos, est liée à
la différence de concentration en ions entre les deux milieux.
Lorsqu'un influx nerveux se déplace le long de la membrane
plasmique, la différence de potentiel transmembranaire
varie brusquement; l'intérieur de la cellule devient positif
(+30 millivolts) pendant quelques millisecondes puis reprend sa
valeur de repos de -70 millivolts. Cette brusque inversion est
appelée potentiel d'action. C'est la manifestation électrique
de l'influx nerveux. La conductibilité d'un neurone est
sa capacité de transmettre l'influx nerveux très
rapidement et sur de longues distances .
4.2. Développement embryologique
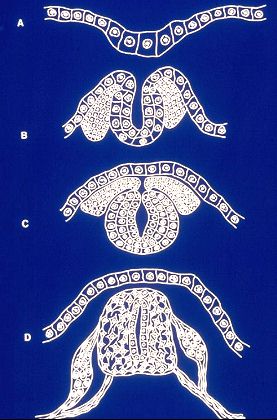
 Le système nerveux se forme à partir d'une bande
ectoblastique médiane et dorsale, le neurectoblaste. Cette
bande s'épaissit et devient la plaque neurale (A). Les
cellules de la plaque s'enfoncent et la plaque neurale devient
gouttière neurale (B). Enfin la gouttière se ferme
pour isoler le tube neural (C); qui est à l'origine du
système nerveux central. Aux bords de la gouttière
naissent des bourgeons cellulaires, les crêtes neurales,
qui glissent sous l'ectoblaste dans le mésenchyme. D'elles
dérivent entre autres toutes les formations qui appartiennent
au système nerveux périphérique.
Le système nerveux se forme à partir d'une bande
ectoblastique médiane et dorsale, le neurectoblaste. Cette
bande s'épaissit et devient la plaque neurale (A). Les
cellules de la plaque s'enfoncent et la plaque neurale devient
gouttière neurale (B). Enfin la gouttière se ferme
pour isoler le tube neural (C); qui est à l'origine du
système nerveux central. Aux bords de la gouttière
naissent des bourgeons cellulaires, les crêtes neurales,
qui glissent sous l'ectoblaste dans le mésenchyme. D'elles
dérivent entre autres toutes les formations qui appartiennent
au système nerveux périphérique.
4.3. Neurone
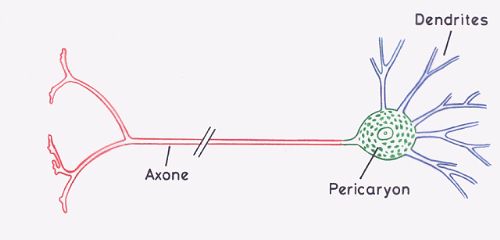
 Le neurone est la cellule qui élabore, modifie et transmet
l'influx nerveux. Il possède un corps cellulaire ou péricaryon
et des prolongements appelés neurites ou fibres nerveuses.
Le péricaryon, représenté en vert, est constitué
du noyau et du cytoplasme qui l'entoure. Les neurites sont de
deux types : les dendrites, représentées en bleu,
et l'axone, représenté en rouge. Les synapses sont
les sites spécialisés de communication entre des
neurones ou entre des neurones et d'autres cellules.
Le neurone est la cellule qui élabore, modifie et transmet
l'influx nerveux. Il possède un corps cellulaire ou péricaryon
et des prolongements appelés neurites ou fibres nerveuses.
Le péricaryon, représenté en vert, est constitué
du noyau et du cytoplasme qui l'entoure. Les neurites sont de
deux types : les dendrites, représentées en bleu,
et l'axone, représenté en rouge. Les synapses sont
les sites spécialisés de communication entre des
neurones ou entre des neurones et d'autres cellules.
La taille et la forme du péricaryon ainsi que le nombre
et le mode de branchement des prolongements sont très variables.
L'aspect des cellules nerveuses dépend aussi des techniques
utilisées pour les mettre en évidence. Voici quelques
neurones de la moelle épinière observés après
une imprégnation à l'argent. Cette technique met
en évidence les faisceaux de neurofilaments qui remplissent
le cytoplasme du péricaryon et se prolongent dans les neurites,
jusque dans leurs plus fines ramifications. L'imprégnation
à l'argent est ainsi une bonne méthode pour identifier
les prolongements cellulaires.
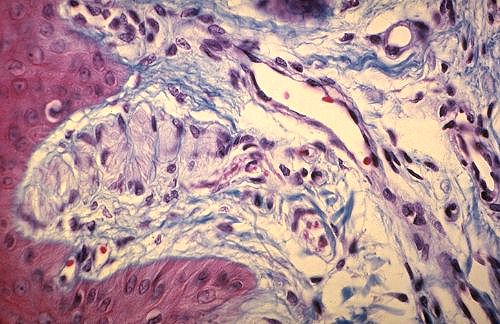
Les mêmes neurones sont photographiés ici après
une coloration trichromique classique. Les péricaryons
de la moëlle épinière ont une forme anguleuse,
polygonale. Chaque angle donne naissance à un neurite.
Le noyau, volumineux et clair, contient peu d'hétérochromatine
et de gros nucléoles. Dans le cytoplasme se trouvent des
amas basophiles, les blocs de Nissl. Entre les neurones sont entremêlés
de nombreux neurites.
 Dans cette micrographie électronique, la composition des
blocs de Nissl est évidente : ce sont des saccules de réticulum
endoplasmique rugueux empilés et parallèles les
uns aux autres. Les autres constituants de la cellule ne sont
pas visibles dans cette image. Le noyau est sphérique et
souvent central; sa chromatine est diffuse et le gros nucléole
est bien visible. Le cytosquelette est très développé;
les neurofilaments et les microtubules traversent le corps cellulaire
en passant d'un prolongement à l'autre. L'appareil de Golgi
est aggloméré dans la région juxtanucléaire.
Les mitochondries sont petites. Le cytoplasme contient encore
des lysosomes, parfois des gouttelettes lipidiques et des pigments.
Dans cette micrographie électronique, la composition des
blocs de Nissl est évidente : ce sont des saccules de réticulum
endoplasmique rugueux empilés et parallèles les
uns aux autres. Les autres constituants de la cellule ne sont
pas visibles dans cette image. Le noyau est sphérique et
souvent central; sa chromatine est diffuse et le gros nucléole
est bien visible. Le cytosquelette est très développé;
les neurofilaments et les microtubules traversent le corps cellulaire
en passant d'un prolongement à l'autre. L'appareil de Golgi
est aggloméré dans la région juxtanucléaire.
Les mitochondries sont petites. Le cytoplasme contient encore
des lysosomes, parfois des gouttelettes lipidiques et des pigments.
Les neurites, axones et dendrites, sont les prolongements des
neurones. Le cytoplasme clair est limité par la membrane
plasmique. Il contient des microtubules, des faisceaux de neurofilaments,
de nombreuses mitochondries et du réticulum endoplasmique
lisse. Chaque neurite est entouré par le cytoplasme d'une
cellule gliale, la cellule de Schwann, que nous décrirons
plus loin.
Les neurites sont impliqués dans la conduction de l'influx
nerveux. L'influx est induit à l'extrémité
de la dendrite; il se déplace le long de celle-ci pour
rejoindre le péricaryon. Il quitte le péricaryon
par l'axone et se déplace vers l'extrémité
synaptique de l'axone. La conduction est donc centripète
dans la dendrite et centrifuge dans l'axone.
Il existe plusieurs différences morphologiques entre
les dendrites et les axones. Les dendrites sont le plus souvent
nombreuses et très ramifiées; les collatérales
s'en détachent à angle aigu et dessinent une figure
spatiale souvent caractéristique d'un type de neurone.
Certaines dendrites sont hérissées d'aspérités
ou épines dendritiques, constituées d'un pédoncule
et d'une dilatation terminale. Les dendrites contiennent des microtubules
et des neurofilaments, du réticulum endoplasmique lisse
et rugueux, des ribosomes libres et des mitochondries.
L'axone est unique. Il naît d'un cône d'émergence,
dont la partie initiale a l'aspect cytoplasmique du péricaryon
et dont la partie distale est pourvue d'une membrane plasmique
épaissie. Ses ramifications sont rares; elles se détachent
à angle droit et leur extrémité synaptique
est dilatée en bouton. La paroi de l'axone est lisse. Comme
la dendrite, il contient des microtubules, des neurofilaments,
du réticulum endoplasmique lisse et de nombreuses mitochondries.
Mais, il est dépourvu de ribosomes libres et de réticulum
endoplasmique rugueux; sa caractéristique principale est
la présence de vésicules qui s'accumulent dans les
boutons terminaux où elles portent le nom de vésicules
synaptiques.
La synapse est le relais qui assure la transmission de l'influx
nerveux d'un neurone à l'autre ou d'un neurone à
une cellule effectrice, musculaire par exemple. Elle comprend
un élément présynaptique et un élément
postsynaptique séparés par une fente synaptique
d'environ 20 à 30 nm. L'élément présynaptique
ou "bouton synaptique" est toujours l'extrémité
renflée d'un axone. lI contient de nombreuses vésicules
synaptiques. L'élément postsynaptique est une portion
spécialisée de la membrane plasmique d'une cellule
effectrice ou d'un neurone.
Lorsque les synapses intéressent deux neurones, on les
classe d'après leur position sur le neurone postsynaptique.
On distingue ainsi les synapses axo-dendritiques, axo-somatiques
et axo-axoniques.
La plupart des synapses sont dites chimiques parce qu'elles impliquent
la libération d'un médiateur, appelé neurotransmetteur,
contenu dans les vésicules synaptiques. Elles sont polarisées
: l'influx passe toujours de l'élément présynaptique
à l'élément postsynaptique. Lors de l'influx,
les vésicules fusionnent avec la paroi du bouton; le neurotransmetteur
libéré diffuse dans la fente et se fixe sur un récepteur
spécifique de la membrane postsynaptique. La liaison du
neurotransmetteur au récepteur membranaire induit la stimulation
ou l'inhibition du second neurone ou de la cellule effectrice.
Le neurotransmetteur est ensuite détruit ou repris par
endocytose dans le bouton présynaptique.
4.4. Architecture du système nerveux périphérique
Le système nerveux périphérique comprend
les ganglions, les nerfs et les terminaisons nerveuses.
4.4.1. Ganglions
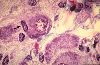 Les corps cellulaires des neurones dans le système nerveux
périphérique se trouvent uniquement dans les ganglions;
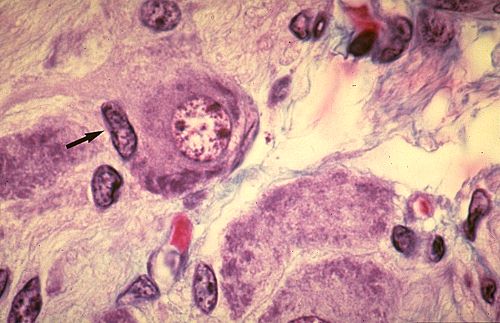
on les appelle cellules ganglionnaires. En voici une. Elle est
volumineuse et possède tous les caractères que nous
venons de décrire; son noyau est central et clair et le
nucléole est bien visible. Son cytoplasme abondant est
basophile et contient des blocs de Nissl. La cellule ganglionnaire
est toujours entourée de cellules gliales, appelées
cellules capsulaires. La cellule capsulaire est petite. Son noyau,
désigné ici par une flèche, est ovoïde
et dense. Son cytoplasme est si mince qu'il n'est généralement
pas visible en microscopie optique.
Les corps cellulaires des neurones dans le système nerveux
périphérique se trouvent uniquement dans les ganglions;
on les appelle cellules ganglionnaires. En voici une. Elle est
volumineuse et possède tous les caractères que nous
venons de décrire; son noyau est central et clair et le
nucléole est bien visible. Son cytoplasme abondant est
basophile et contient des blocs de Nissl. La cellule ganglionnaire
est toujours entourée de cellules gliales, appelées
cellules capsulaires. La cellule capsulaire est petite. Son noyau,
désigné ici par une flèche, est ovoïde
et dense. Son cytoplasme est si mince qu'il n'est généralement
pas visible en microscopie optique.
Tout ganglion nerveux se compose donc de péricaryons et
des cellules capsulaires qui les enveloppent étroitement.
Mais il contient aussi le début des neurites qui naissent
de ces cellules ganglionnaires et qui sont enveloppés de
leurs propres cellules gliales ou cellules de Schwann; d'autres
fibres nerveuses le traversent ou s'y terminent. Parmi les cellules
nerveuses et les cellules gliales se trouvent des capillaires
sanguins et un peu de tissu conjonctif. Le ganglion est enveloppé
d'une capsule conjonctive dense qui se prolonge dans la gaine
conjonctive des nerfs qui y aboutissent ou le quittent. Ces nerfs
ne sont pas visibles dans l'image.
Parmi les ganglions, on distingue les ganglions rachidiens
et les ganglions autonomiques. Les ganglions rachidiens sont
situés sur la racine dorsale des nerfs rachidiens, issus
de la moelle épinière. Ils contiennent des neurones
sensitifs, unipolaires, ne possédant qu'un seul prolongement
qui se divise à distance du corps cellulaire en un axone
et une dendrite.
Les ganglions autonomiques appartiennent au système nerveux
autonomique qui règle les fonctions végétatives,
indépendantes de la volonté. Les neurones y sont
moteurs et multipolaires. Parmi les ganglions autonomiques, on
distingue les ganglions prévertébraux disposés
en chaîne de chaque côté de la colonne vertébrale
et les ganglions situés à distance de ces chaînes,
dont les ganglions viscéraux qui se trouvent dans la paroi
des viscères.
4.4.2. Nerf
Le nerf est un ensemble de complexes axo-schwanniens, groupés
en un ou plusieurs faisceaux. Chaque complexe est l'association
d'un neurite, axone ou dendrite, avec des cellules gliales, les
cellules de Schwann, qui lui fournissent éventuellement
une gaine de myéline.
Dans le complexe axo-schwannien, le neurite est entièrement
logé dans une invagination de la membrane plasmique de
la cellule de Schwann de telle sorte qu'il paraît, en coupe,
incorporé dans son cytoplasme. Lorsque la membrane invaginée
de la cellule de Schwann n'est pas modifiée, le neurite
est dit amyélinisé et la même cellule peut
alors, comme ici, en contenir plusieurs. Les fibres collagènes
qui l'entourent appartiennent à l'endonèvre.
Dans d'autres complexes axo-schwanniens, le neurite est totalement
enveloppé d'une épaisse couche de myéline.
ll n'existe alors qu'un seul neurite par cellule de Schwann. Cette
gaine de myéline provient de l'enroulement de la membrane
plasmique invaginée de la cellule de Schwann autour du
neurite et de la fusion des membranes de spires accolées.
L'enroulement et les fusions membranaires expliquent la structure
de la gaine et sa striation très particulière.
La striation de la myéline s'explique plus facilement dans
le schéma d'une fibre myélinisée en coupe
longitudinale, à gauche, et en coupe transversale, à
droite. Le noyau et le cytoplasme de la cellule de Schwann sont
tassés dans la partie la plus externe du complexe axo-schwannien.
Dans toute son épaisseur, la myéline est caractérisée
(coupe B) par l'alternance de lignes épaisses, dites périodiques,
et de lignes minces, dites intrapériodiques. Parfois (coupe
A), en quelques endroits, de petites restes cytoplasmiques interrompent
la ligne périodique; ce sont les incisures de Schmidt-Lanterman
qui traduisent un défaut dans la fusion des membranes.
La gaine de myéline est discontinue : entre deux cellules
de Schwann (coupe C), le neurite est uniquement enveloppé
par le glycocalix. Ces intervalles sont les noeuds de Ranvier.
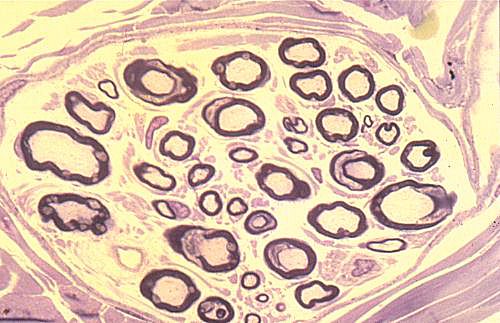
 Voici la section transversale d'un faisceau nerveux dans une coupe
semi-fine colorée au bleu de toluidine. Les neurites sont
les plages claires plus ou moins rondes. Ils sont myélinisés
: la gaine de myéline est un anneau coloré en bleu
foncé, autour de ces plages. Les incisures de Schmidt-Lanterman,
très nombreuses, sont de petites taches claires dans les
anneaux de myéline.
Voici la section transversale d'un faisceau nerveux dans une coupe
semi-fine colorée au bleu de toluidine. Les neurites sont
les plages claires plus ou moins rondes. Ils sont myélinisés
: la gaine de myéline est un anneau coloré en bleu
foncé, autour de ces plages. Les incisures de Schmidt-Lanterman,
très nombreuses, sont de petites taches claires dans les
anneaux de myéline.
Puisqu'elle provient de la fusion de membranes cellulaires, la
myéline est composée en grande partie de phospholipides.
La majorité d'entre eux sont extraits par les solvants
utilisés pour l'enrobage à la paraffine. Les fibres
nerveuses, vues en coupe longitudinale, sont allongées
parallèlement les unes aux autres. Lorsque la coupe passe
dans l'axe d'un complexe axo-schwannien, on peut observer le neurite
homogène et grisâtre. Il est entouré de part
et d'autre par une bande claire qui contient un matériel
granulaire rouge, représentant les restes protéiques
de la myéline. Chaque complexe est isolé de ses
voisins par une lamelle de tissu conjonctif. Les noyaux appliqués
contre les complexes appartiennent aux cellules de Schwann dont
le cytoplasme n'est pas visible. Au niveau des noeuds de Ranvier,
la gaine de myéline est interrompue par un repli et le
neurite y est plus coloré.
Dans une coupe à congélation, la myéline
est mise en évidence par des techniques histochimiques.
Sa richesse en sphingolipides est décelée par le
bleu de Nil qui lui donne, dans cette coupe transversale de nerf,
l'aspect d'un anneau bleu. Les neurites, non colorés, sont
au centre des anneaux.
 Dans un nerf en coupe transversale, les complexes axo-schwanniens,
petites plages claires et rondes, sont séparés les
uns des autres par l'endonèvre coloré en bleu. Ils
sont groupés en faisceaux. La plupart des noyaux à
l'intérieur des faisceaux appartiennent aux cellules de
Schwann. Chaque faisceau est enveloppé par du périnèvre,
indiqué par des flèches. L'épinèvre,
composé de tissu conjonctif, enveloppe le nerf et relie
les faisceaux entre eux.
Dans un nerf en coupe transversale, les complexes axo-schwanniens,
petites plages claires et rondes, sont séparés les
uns des autres par l'endonèvre coloré en bleu. Ils
sont groupés en faisceaux. La plupart des noyaux à
l'intérieur des faisceaux appartiennent aux cellules de
Schwann. Chaque faisceau est enveloppé par du périnèvre,
indiqué par des flèches. L'épinèvre,
composé de tissu conjonctif, enveloppe le nerf et relie
les faisceaux entre eux.
L'endonèvre est un tissu conjonctif où les fibres
collagènes sont disposées longitudinalement entre
les complexes axo-schwanniens. Il contient des fibroblastes (F)
et, non visibles dans la région choisie, des capillaires.
Le périnèvre, qui délimite chaque faisceau,
est un tissu lamellaire, dont une cellule est désignée
par une flèche.
Chaque lame périneurale est formée d'une seule assise
cellulaire soulignée de chaque côté par un
épais glycocalix et séparée de ses voisines
par des fibres collagènes. Les cellules périneurales
sont fines et ramifiées; leurs extémités
sont accolées les unes aux autres.
Le nerf est donc une association de complexes axo-schwanniens,
myélinisés ou non, rassemblés en un ou plusieurs
faisceaux. Les complexes axo-schwanniens sont séparés
les uns des autres par l'endonèvre, fine lame de tissu
conjonctif. Chaque faisceau est limité par le périnèvre,
formé de lames cellulaires et de fibres de collagène.
Les faisceaux adhèrent entre eux et au tisu avoisinant
par l'épinèvre composé de tissu conjonctif
dense.
4.4.3. Terminaisons nerveuses
La terminaison nerveuse est l'extrémité d'un
neurite dans un organe périphérique. Elle est sensitive
lorsqu'elle est le début d'une dendrite. Elle est motrice
lorsqu'elle est la fin d'un axone.
La terminaison sensitive est un récepteur sensoriel qui
convertit les stimuli provenant du milieu intérieur ou
extérieur en influx nerveux qui sont transmis aux péricaryons.
Il existe trois groupes de terminaisons sensitives: les
terminaisons libres ou nues, les terminaisons encapsulées
sans support et les terminaisons encapsulées avec support.
Dans certains organes des sens, les terminaisons sensitives forment
des structures neuroépithéliales.
La terminaison nerveuse sensitive libre (1) est l'extrémité
d'une dendrite dépourvue de toute gaine et ramifiée
en plusieurs branches. Elle se glisse entre les cellules, notamment
dans les épithéliums, et recueille de façon
peu sélective les stimuli mécaniques, thermiques
ou algésiques. La terminaison encapsulée sans support
(2) est totalement séparée du tissu environnant
par une capsule composée de cellules périneurales
et de tissu conjonctif. Nous en verrons deux exemples, le corpuscule
de Meissner et le corpuscule de Vater Paccini. Dans la terminaison
encapsulée avec support (3), la dendrite est enroulée
autour d'éléments appartenant au tissu où
elle se trouve. Ces éléments, cellules musculaires
dans le fuseau neuro-musculaire ou fibres conjonctives dans l'organe
tendineux de Golgi, constituent le support. L'extrémité
dendritique et le support sont enveloppés dans la capsule
formée par le périnèvre et un reste d'endonèvre.
Le fuseau neuro-musculaire sera décrit plus en détail
dans le chapitre sur le muscle.
Le corpuscule de Meissner (A) est situé sous l'épithélium
épidermique et est impliqué dans la sensibilité
tactile fine. Il est piriforme et mesure 120 µm de long et
60 µm de large. L'extrémité dendritique, dépourvue
de gaine de myéline, de gaine de Schwann et d'endonèvre,
est pelotonnée. La capsule périneurale est très
mince. Le corpuscule de Vater-Paccini (B) est un récepteur
de la pression profonde, notamment dans le derme. Il a la forme
d'une ampoule et peut mesurer 4 mm de long et 2 mm de large. Sa
capsule, très épaisse, est constituée d'une
série de lames périneurales disposées en
bulbe d'oignon. Les lames sont séparées par du tissu
conjonctif vascularisé. Le centre du corpuscule est occupé
par une ou plusieurs extrémités dendritiques amyélinisées.
La gaine de myéline commence dès la sortie du corpuscule.
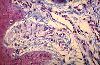 Le corpuscule de Meissner est toujours situé dans une papille
dermique.
Le corpuscule de Meissner est toujours situé dans une papille
dermique.
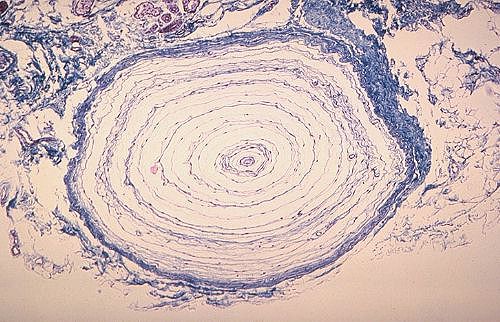
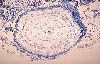 Voici un corpuscule de Vater-Paccini dans le derme profond.
Voici un corpuscule de Vater-Paccini dans le derme profond.
La structure neuroépithéliale est l'association
de dendrites et de cellules localisées dans un épithélium,
originaires des crêtes ganglionnaires et qui reçoivent
le stimulus. On observe ce genre de structure dans les fosses
nasales, l'oreille, la peau et la langue. Le bourgeon gustatif,
par exemple, est l'organe neuroépithélial du goût.
Il a la forme d'un tonnelet au sein de l'épithélium
épidermoïde qui recouvre la langue. Sa base est en
contact avec le chorion lâche. Son sommet arrive à
la surface de l'épithélium. Il contient une petite
cavité, le canal gustatif.
Ce schéma de bourgeon gustatif montre ses différents
types de cellules. Certaines, notamment celles de type III, ont
de courtes microvillosités qui baignent dans le canal gustatif.
Au niveau de zones membranaires épaissies, elles sont en
rapport étroit avec des fibres nerveuses sensitives, teintées
en bleu dans le schéma. Leur cytoplasme contient de nombreuses
petites vésicules, analogues aux vésicules synaptiques
et concentrées près de ces zones. Les substances
sapides se lient à des récepteurs membranaires des
microvillosités; cette liaison provoque l'exocytose des
vésicules et donc la stimulation nerveuse.
Les terminaisons motrices ou effectrices sont les extrémités
des axones. Elles provoquent ou modulent la contraction des muscles
ou règlent la sécrétion des glandes. Dans
les glandes, l'extrémité nue et dilatée de
l'axone s'insère dans une invagination de la membrane plasmique
de la cellule glandulaire. Dans les muscles lisse et cardiaque,
l'extrémité nue se termine à distance des
cellules musculaires. Dans le muscle squelettique, elle entre
dans la constitution d'une plaque motrice que nous étudierons
en détail avec les autres jonctions neuro-musculaires.
Leçon suivante: 4.5. Architecture du système
nerveux central
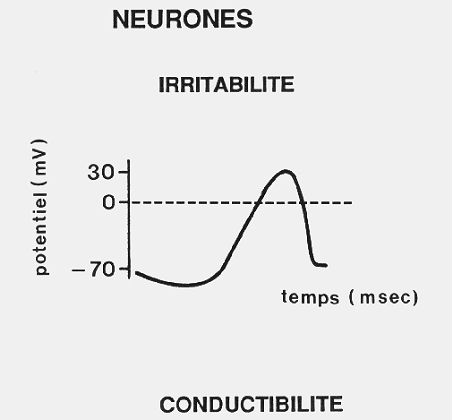 Les fonctions du système nerveux sont basées sur
deux propriétés cellulaires, l'irritabilité
et la conductibilité, particulièrement
développées dans le neurone. Son irritabilité
est sa capacité de réagir aux variations du milieu
extracellulaire en modifiant les propriétés électriques
de sa membrane. Lorsqu'aucun influx nerveux n'est transmis, le
potentiel électrique à l'intérieur de la
cellule est négatif d'environ -70 millivolts par rapport
à celui de l'extérieur; cette différence
de potentiel, ou potentiel de repos, est liée à
la différence de concentration en ions entre les deux milieux.
Lorsqu'un influx nerveux se déplace le long de la membrane
plasmique, la différence de potentiel transmembranaire
varie brusquement; l'intérieur de la cellule devient positif
(+30 millivolts) pendant quelques millisecondes puis reprend sa
valeur de repos de -70 millivolts. Cette brusque inversion est
appelée potentiel d'action. C'est la manifestation électrique
de l'influx nerveux. La conductibilité d'un neurone est
sa capacité de transmettre l'influx nerveux très
rapidement et sur de longues distances .
Les fonctions du système nerveux sont basées sur
deux propriétés cellulaires, l'irritabilité
et la conductibilité, particulièrement
développées dans le neurone. Son irritabilité
est sa capacité de réagir aux variations du milieu
extracellulaire en modifiant les propriétés électriques
de sa membrane. Lorsqu'aucun influx nerveux n'est transmis, le
potentiel électrique à l'intérieur de la
cellule est négatif d'environ -70 millivolts par rapport
à celui de l'extérieur; cette différence
de potentiel, ou potentiel de repos, est liée à
la différence de concentration en ions entre les deux milieux.
Lorsqu'un influx nerveux se déplace le long de la membrane
plasmique, la différence de potentiel transmembranaire
varie brusquement; l'intérieur de la cellule devient positif
(+30 millivolts) pendant quelques millisecondes puis reprend sa
valeur de repos de -70 millivolts. Cette brusque inversion est
appelée potentiel d'action. C'est la manifestation électrique
de l'influx nerveux. La conductibilité d'un neurone est
sa capacité de transmettre l'influx nerveux très
rapidement et sur de longues distances .