Les organes lymphoïdes (cours 3)
 La rate (1) est
un organe ovoïde situé dans la loge splénique
de l'hypochondre gauche. Elle est entièrement enveloppée
par le péritoine sauf au niveau de son bord interne
ou hile qui donne insertion aux ligaments gastrosplénique
et pancréatico-splénique. Par l'intermédiaire
du péritoine, la rate est en rapport avec le diaphragme
en haut et en arrière, l'estomac (2) en avant, l'angle
colique gauche et le rein gauche (3) en bas. La taille de la rate
est très variable. Normalement, ses diamètres ne
dépassent pas 13 cm en longueur et 8 cm en largeur et elle
ne déborde pas le rebord costal.
La rate (1) est
un organe ovoïde situé dans la loge splénique
de l'hypochondre gauche. Elle est entièrement enveloppée
par le péritoine sauf au niveau de son bord interne
ou hile qui donne insertion aux ligaments gastrosplénique
et pancréatico-splénique. Par l'intermédiaire
du péritoine, la rate est en rapport avec le diaphragme
en haut et en arrière, l'estomac (2) en avant, l'angle
colique gauche et le rein gauche (3) en bas. La taille de la rate
est très variable. Normalement, ses diamètres ne
dépassent pas 13 cm en longueur et 8 cm en largeur et elle
ne déborde pas le rebord costal.
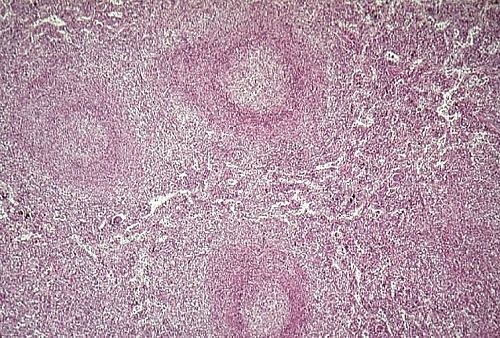
Dans une tranche macroscopique
on peut déjà distinguer les différentes parties
de la rate. Les bords de la tranche sont délimités
par une épaisse membrane, la capsule splénique.
La surface de la tranche est parsemée de points blancs,
la pulpe blanche. Tout ce qui se trouve entre ces points
blancs est la pulpe rouge, habituellement pâteuse
lorsque la rate est fraîchement prélevée .
Elle est rouge parce que gorgée de sang.
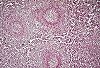 Un examen microscopique
à faible grossissement permet de préciser la nature
des pulpes spléniques. La pulpe blanche comprend de nombreux
amas lymphoïdes. La pulpe rouge est un réseau
d'espaces entre les formations lymphoïdes. Chaque pulpe
est composée de deux structures qui sont représentées
dans le schéma suivant.
Un examen microscopique
à faible grossissement permet de préciser la nature
des pulpes spléniques. La pulpe blanche comprend de nombreux
amas lymphoïdes. La pulpe rouge est un réseau
d'espaces entre les formations lymphoïdes. Chaque pulpe
est composée de deux structures qui sont représentées
dans le schéma suivant.
De la capsule splénique (1) naissent des travées
importantes (2) qui cloisonnent incomplètement le parenchyme
de l'organe. Dans ces travées se trouvent les gros vaisseaux.
La pulpe blanche est constituée de manchons périartériolaires
(3) et de centres germinatifs (4). La pulpe rouge
comprend de larges vaisseaux, les sinusoïdes veineux
(5) et les cordons de Billroth (6) qui sont des espaces
réticulaires entre les sinusoïdes veineux.
La capsule splénique
est épaisse et dense. Sa face externe est couverte par
un mésothélium, le feuillet péritonéal
viscéral. Sa composition varie selon les espèces.
Chez l'homme et les rongeurs, les fibres musculaires lisses sont
rares et la capsule est essentiellement conjonctive.
Dans d'autres espèces, le chat, le chien, mais particulièrement
les grands carnivores sauvages, la capsule est surtout musculaire.
Lors d'un effort brusque et important, les fibres musculaires
lisses se contractent, les globules rouges qui stagnent dans la
rate, sont expulsés dans la circulation.
Les travées qui proviennent de cette capsule ont naturellement
la même structure, mais sont plus riches en fibres élastiques.
Voici la coupe transversale
d'une fine travée presqu'exclusivement conjonctive.
L'extrémité d'une travée
est réduite à quelques fibres musculaires
coupées ici longitudinalement. Les divers aspects de ces
travées ne peuvent être confondus avec les autres
structures spléniques.
Les manchons périartériolaires
sont des amas de lymphocytes qui enveloppent de petites
branches artérielles, les artères centrales ou
nodulaires. Ces manchons sont rarement observés en
coupe longitudinale. Ils le sont plus souvent en coupe transversale.
C'est pour cette raison qu'ils ont été d'abord décrits
comme des nodules, les nodules de Malpighi, spécifiques
de la rate parce que traversés par un rameau artériel.
Dans cette préparation qui provient d'un animal perfusé,
l'artère centrale est dilatée et ne contient pas
de globules rouges.
Dans ces manchons,
les nombreux petits et moyens lymphocytes sont emprisonnés
dans les mailles de tissu réticulé. Les macrophages,
les plasmocytes et les globules rouges sont rares. Comme dans
le ganglion, les cellules réticulées forment autour
du manchon une nappe cellulaire qui enveloppe étroitement
la masse lymphoïde.
Répartis dans toute la rate mais toujours en rapport avec
les manchons périartériolaires, les centres germinatifs
ont la même fonction et le même aspect que dans les
autres organes lymphoïdes.
On leur décrit donc une zone plus sombre,
riche en immunoblastes, en macrophages et en mitoses. La
zone plus claire est composée surtout de lymphocytes
et de cellules dendritiques. Elle est enveloppée d'une
coiffe très dense de petits lymphocytes.
Dans la pulpe rouge,
les sinusoïdes veineux sont identifiables parce que
ce sont de larges espaces vasculaires qui apparaissent
plus clairs que les structures voisines. Ils contiennent de nombreux
éléments sanguins et sont délimités
par un endothélium discontinu.
Cette discontinuité est bien visible lorsque l'on peut
examiner la face externe d'un sinusoïde
au microscope électronique à balayage. La paroi
est percée d'orifices délimités par des cellules
étoilées anastomosées entre elles. A
travers les orifices on aperçoit la cavité du sinusoïde
et les globules rouges, en forme de disque, qu'il contient.
Grâce à ces orifices, la paroi du sinusoïde
est perméable à tous les éléments
figurés du sang. Ici, la fixation a saisi un lymphocyte
qui s'étire entre les cellules endothéliales. Bien
sûr, on ne peut décider sur la base de telles images
si ces cellules quittent le sinusoïde ou y pénètrent.
Les cordons de Billroth
sont situés entre les sinusoïdes veineux et entre
ceux-ci et les autres structures de la rate. Leurs mailles contiennent
tous les éléments sanguins. Elles sont particulièrement
riches en macrophages reconnaissables sans doute à
leur taille, à l'excentricité de leur noyau ou à
l'irrégularité de leurs contours, mais surtout à
leurs vacuoles cytoplasmiques ou, comme ici, aux inclusions
pigmentaires brunâtres.
Ces mailles sont faites de cellules réticulées
comme celles des sinus médullaires du ganglion. Cette image,
prise au microscope électronique à balayage, montre
comment ces cellules s'articulent pour former un véritable
labyrinthe.
La distinction entre sinusoïdes veineux et cordon de Billroth
est très nette lorsque la réticuline
est mise en évidence. Dans le tissu réticulé
des cordons, les fibres forment un réseau dont les mailles
sont de taille variable. Le squelette fibreux des sinusoïdes,
coupés longitudinalement, apparaît comme une suite
de points foncés, parce que les fibres de réticuline
sont concentriques par rapport à la lumière du
vaisseau.
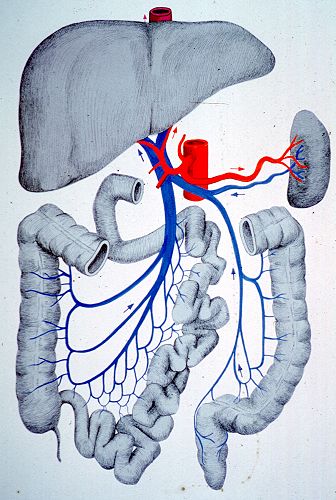
 La vascularisation lymphatique de la rate n'est pas connue. Sa
vascularisation sanguine
est importante car c'est d'elle que dépendent en grande
partie les fonctions spléniques. Une seule artère
assure toute la vascularisation; c'est l'artère splénique,
branche du tronc coeliaque qui aboutit au hile de l'organe et
s'y divise en plusieurs branches. Le retour veineux est assuré
par la veine splénique qui reçoit la petite
veine mésentérique puis s'unit à la grande
veine mésentérique derrière la tête
du pancréas pour former la veine porte. Tout le sang en
provenance de la rate est donc destiné au foie.
La vascularisation lymphatique de la rate n'est pas connue. Sa
vascularisation sanguine
est importante car c'est d'elle que dépendent en grande
partie les fonctions spléniques. Une seule artère
assure toute la vascularisation; c'est l'artère splénique,
branche du tronc coeliaque qui aboutit au hile de l'organe et
s'y divise en plusieurs branches. Le retour veineux est assuré
par la veine splénique qui reçoit la petite
veine mésentérique puis s'unit à la grande
veine mésentérique derrière la tête
du pancréas pour former la veine porte. Tout le sang en
provenance de la rate est donc destiné au foie.
La circulation intrasplénique
est complexe et toujours discutée. Chaque branche de l'artère
splénique (1) se ramifie dans les travées
(2). A chaque division, les artérioles diminuent de calibre
et lorsque leur diamètre ne dépasse plus deux dixièmes
de millimètre, elles quittent les travées. On les
appelle dès lors les artères centrales (3)
parce qu'elles sont immédiatement enveloppées par
les manchons périartériolaires de la pulpe blanche.
Ces artères centrales se ramifient un petit nombre de fois.
Lorsque leur diamètre est d'environ 50 microns, elles se
divisent en un pinceau de petites branches, les artères
pénicillées (4) qui quittent progressivement
la pulpe blanche pour pénétrer dans la pulpe rouge.
Elles donnent naissance à de courts capillaires
(5). A partir d'ici les opinions divergent. Pour les tenants de
la circulation ouverte (A), ces capillaires s'ouvrent dans
les espaces réticulés des cordons de Billroth
(6). Ceux- ci sont drainés par les sinusoïdes
(7) qui s'unissent pour former les veines septales (8).
Toutes convergent vers le hile où elles se jettent dans
une branche de la veine splénique (9). Pour les
partisans de la circulation fermée (B), les capillaires
des artères pénicillées ne s'ouvrent pas
dans les espaces réticulés mais sont en continuité
avec les sinusoïdes veineux. Les deux types de circulation
pourraient coexister ou alterner selon l'état physiologique
de la rate.
L'artère septale
se reconnaît facilement. Sa lumière est délimitée
par un endothélium pavimenteux simple. Sa média
musculeuse est épaisse. Les fibres musculaires lisses,
disposées en plusieurs couches, sont ici coupées
longitudinalement parce que la coupe dans l'artère est
transversale. L'adventice est réduite au peu de tissu conjonctif
qui appartient à la travée
Les artères centrales
sont beaucoup plus petites. L'endothélium est continu.
La média musculeuse est réduite. L'adventice est
constituée uniquement des lymphocytes appartenant
aux gaines périartériolaires. Le terme "artère
centrale" est un peu abusif car elles sont rarement situées
au centre de leur manchon qui est d'ailleurs fréquemment
déformé par l'apparition des centres germinatifs.
Ceux-ci ne possèdent pas d'artère.
Les plus petites artères centrales se divisent en artères
pénicillées. Voici cette division en coupe transversale.
Les artères pénicillées sont très
petites. Le diamètre de leur lumière n'excède
pas celui d'un globule rouge. Leur média est réduite
à une seule couche de muscle lisse et est parfois enveloppée
d'une mince adventice conjonctive.
Voici l'artère pénicillée
en coupe longitudinale, telle qu'on l'observe le plus fréquemment.
Sa lumière est délimitée par les cellules
endothéliales dont on voit les noyaux allongés.
La média est représentée par les quelques
noyaux plus épais situés en dehors de l'endothélium.
Cette média devient de moins en moins importante à
mesure que l'on se rapproche de la pulpe rouge où l'artère
pénicillée se résout en très courts
capillaires.
Dans certaines espèces, en particulier les carnivores comme
le chien et la chat, ces capillaires de la pulpe rouge ont une
enveloppe spéciale, la housse de Schweigger-Seidel
composée d'un amas de cellules dites épithélioïdes
...
Cette housse peut envelopper un capillaire ou l'ensemble des capillaires
issus d'une artère et est fréquemment infiltrée
de globules rouges. Sa signification n'est pas connue. Elle n'existe
ni chez l'homme ni chez les rongeurs.
La transition entre les branches des artères pénicillées
et le reste de la circulation est rarement observée; le
choix entre la circulation ouverte et la circulation fermée
est donc difficile, certaines images plaident cependant en faveur
de la première. Ici, par exemple, l'artère pénicillée
paraît s'ouvrir dans le réticulum d'un cordon de
Billroth.
La même impression se dégage de l'analyse de cette
micrographie, prise
au microscope électronique à balayage.
Les sinusoïdes sont drainés par des veinules qui aboutissent
aux veines septales.
Celles-ci sont larges, souvent gorgées de sang. La lumière
est délimitée par un endothélium. La média
se confond avec le tissu de la travée dans laquelle elle
se trouve. Rappelons que cette travée contient chez certaines
espèces des fibres musculaires lisses dont la disposition
est longitudinale. On ne peut donc confondre ces fibres
musculaires qui accompagnent la veine septale avec l'anneau musculaire
continu d'une artère.
La rate est avant tout un filtre disposé sur la
circulation artérielle. C'est aussi un organe immunitaire
périphérique. Pendant la vie embryonnaire, elle
est hématopoïétique. Elle est parfois
considérée comme une réserve de globules
rouges. Elle peut l'être dans les espèces qui possèdent
une grande rate dont la musculature est développée.
Chez elles, la rate se contracte lors d'un effort puissant. Chez
l'homme cette fonction est très peu importante car le volume
de globules rouges spléniques ne dépasse pas 50
ml. La rate n'est pas indispensable. En cas de splénectomie,
ses fonctions sont reprises par d'autres organes.
Le filtre splénique agit au niveau de la circulation sanguine,
de la même façon que les filtres ganglionnaires agissent
au niveau de la circulation lymphatique.
Mais le rôle principal de cette filtration - et pratiquement
le seul dans les conditions normales - est l'élimination
des globules rouges.
Cette élimination a lieu dans la pulpe rouge. Les
globules rouges
vieillis s'accolent aux macrophages, à cause d'une
modification de leur membrane.
Ils sont ensuite phagocytés
et pendant un temps très bref on peut les reconnaître
entiers dans le cytoplasme des macrophages, mais très rapidement
leur hémoglobine est dégradée.
Cette dégradation libère du fer, partiellement
fixé dans l'hémosidérine. Celle-ci
est un pigment intracellulaire qui apparaît sous forme de
granules brunâtres
de toutes tailles.
Le fer fixé dans l'hémosidérine peut être
coloré par le bleu de Prusse
dans des coupes traitées suivant la technique de Perls,
utilisée en pathologie pour détecter toute surcharge
ferrique. Les noyaux sont ici mis en évidence par le rouge
nucléaire. Une autre fraction du fer est fixée sur
une protéine plasmatique, la transferrine. Elle
servira dans la moelle a synthétiser de la nouvelle hémoglobine.
La portion hémique de l'hémoglobine dégradée
diffuse dans le plasma, arrive au foie, et y est transformée
en bilirubine.
La fonction immunitaire de la rate est la même que celles
des ganglions. Elle intervient donc dans l'immunité
humorale et dans l'immunité cellulaire. Le siège
de cette fonction est la pulpe blanche.
La pulpe blanche
est contrôlée par les organes lymphoïdes centraux.
La partie centrale des manchons périartériolaires
est thymodépendante. La partie périphérique
de ces manchons et les centres germinatifs sont dépendants
de l'homologue de la bourse de Fabricius. Cette fonction
immunitaire comme les autres d'ailleurs n'est pas déterminante.
La splénectomie n'affecte pas l'immunité.
La fonction hématopoïétique de la rate n'existe,
chez l'homme, que pendant la vie embryonnaire. Elle est
normalement reprise par la moelle osseuse peu avant la naissance.
Une recrudescence de l'hématopoïèse splénique
s'observe lors d'une défaillance pathologique de la moelle.
Chez certains rongeurs elle persiste normalement durant la vie
adulte; c'est pourquoi on trouve dans leurs cordons de Billroth
des cellules qui appartiennent à la lignée des granulocytes,
des érythroblastes ou comme ici des mégacaryocytes.
Nous avons terminé ainsi l'étude de la rate. La
leçon suivante est consacrée
aux structures lymphoïdes annexées au tube digestif.
 La rate (1) est
un organe ovoïde situé dans la loge splénique
de l'hypochondre gauche. Elle est entièrement enveloppée
par le péritoine sauf au niveau de son bord interne
ou hile qui donne insertion aux ligaments gastrosplénique
et pancréatico-splénique. Par l'intermédiaire
du péritoine, la rate est en rapport avec le diaphragme
en haut et en arrière, l'estomac (2) en avant, l'angle
colique gauche et le rein gauche (3) en bas. La taille de la rate
est très variable. Normalement, ses diamètres ne
dépassent pas 13 cm en longueur et 8 cm en largeur et elle
ne déborde pas le rebord costal.
La rate (1) est
un organe ovoïde situé dans la loge splénique
de l'hypochondre gauche. Elle est entièrement enveloppée
par le péritoine sauf au niveau de son bord interne
ou hile qui donne insertion aux ligaments gastrosplénique
et pancréatico-splénique. Par l'intermédiaire
du péritoine, la rate est en rapport avec le diaphragme
en haut et en arrière, l'estomac (2) en avant, l'angle
colique gauche et le rein gauche (3) en bas. La taille de la rate
est très variable. Normalement, ses diamètres ne
dépassent pas 13 cm en longueur et 8 cm en largeur et elle
ne déborde pas le rebord costal.