Les organes lymphoïdes (cours 2)
 Les ganglions lymphatiques
sont de petits organes ovoïdes situés sur le
trajet des voies lymphatiques. Les voici tels que les radiologues
les observent après avoir injecté dans un vaisseau
lymphatique une substance opaque aux R.X. Ils sont parfois isolés,
mais le plus souvent groupés là où confluent
les lymphatiques d'un territoire. Les lymphatiques efférents
et les vaisseaux sanguins, abordent ou quittent le ganglion au
niveau du hile.
Les ganglions lymphatiques
sont de petits organes ovoïdes situés sur le
trajet des voies lymphatiques. Les voici tels que les radiologues
les observent après avoir injecté dans un vaisseau
lymphatique une substance opaque aux R.X. Ils sont parfois isolés,
mais le plus souvent groupés là où confluent
les lymphatiques d'un territoire. Les lymphatiques efférents
et les vaisseaux sanguins, abordent ou quittent le ganglion au
niveau du hile.
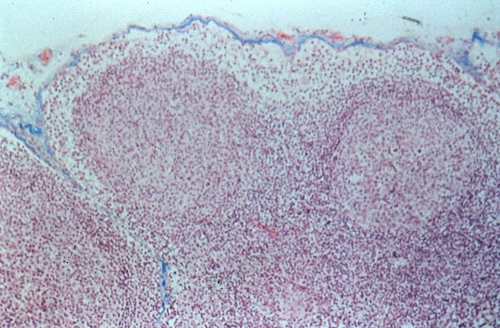
Voici à faible grossissement, l'aspect du ganglion.
Il est enveloppé d'une capsule fibreuse, plus importante
au niveau du hile. Plusieurs travées se détachent
de la face interne de cette capsule et pénètrent
dans la masse du ganglion. Dans cette préparation, elles
sont rares et coupées transversalement.
Le squelette du ganglion est un tissu réticulé.
Celui-ci est infiltré de lymphocytes. Leur répartition
inégale permet de distinguer deux régions. La périphérie
ou cortex ganglionnaire, est une large bande lymphocytaire
très dense et donc très colorée. Elle est
ellemême subdivisée en deux parties : le cortex
externe et le cortex interne. Dans le cortex externe,
on observe des centres germinatifs pâles et de forme
arrondie. Le cortex interne en est dépourvu. La partie
centrale, ou médullaire ganglionnaire est beaucoup
plus claire dans une préparation histologique courante
parce qu'elle est surtout composée de larges espaces réticulés,
les sinus médullaires...
Entre les sinus, se trouvent des expansions du cortex, les prolongements
ou cordons médullaires, anastomosés en réseau.
Il existe entre la capsule et le cortex ganglionnaire, un espace
plus clair, le sinus marginal ou souscapsulaire.
Il communique avec les sinus médullaires par les sinus
intermédiaires qui traversent le cortex.
L'importance relative du cortex et de la médullaire est
très variable. Les ganglions mésentériques
ou lomboaortiques dont le volume est important ont une médullaire
bien développée. Les ganglions jugulaires ont une
médullaire peu importante.
Ce schéma
résume la description précédente. On y retrouve
de dehors en dedans, la capsule fibreuse (1), le sinus
marginal souscapsulaire (2), le cortex ganglionnaire
externe et ses centres germinatifs (3), le cortex ganglionnaire
interne traversé par des septa conjonctifs (4), la
médullaire du ganglion (5) et les prolongements
médullaires du cortex (6).
 La capsule du ganglion
est constituée de tissu conjonctif dense et est
infiltrée, près du hile, de tissu adipeux. Elle
contient des vaisseaux et de rares fibres musculaires lisses autour
des lymphatiques qui la traversent. Les travées qui en
dépendent, cloisonnent incomplètement le parenchyme
ganglionnaire et contiennent des fibres élastiques. L'importance
de ces travées est très variable suivant la localisation,
l'âge et l'espèce.
La capsule du ganglion
est constituée de tissu conjonctif dense et est
infiltrée, près du hile, de tissu adipeux. Elle
contient des vaisseaux et de rares fibres musculaires lisses autour
des lymphatiques qui la traversent. Les travées qui en
dépendent, cloisonnent incomplètement le parenchyme
ganglionnaire et contiennent des fibres élastiques. L'importance
de ces travées est très variable suivant la localisation,
l'âge et l'espèce.
Le tissu conjonctif propre au ganglion est le tissu réticulé
composé de cellules réticulées et
de fibres de réticuline. Ces fibres sont décelées
ici par une imprégnation du tissu à l'argent. Elles
dessinent un réseau dont la forme et l'importance varient
d'une région à l'autre.
Dans le sinus marginal,
les fibres sont épaisses et peu nombreuses. Elles traversent
le sinus et se prolongent dans la capsule, ancrant ainsi le squelette
ganglionnaire au tissu fibreux qui enveloppe le ganglion.
Dans la corticale,
les centres germinatifs sont pratiquement dépourvus de
réticuline. Dans leur périphérie, les fibres
sont épaisses et concentriques. Plus en dehors, elles sont
comparables aux fibres de la médullaire.
Dans la médullaire,
les fibres sont nombreuses et leur enchevêtrement produit
de larges mailles.
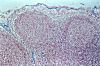 Le cortex ganglionnaire
est habituellement partagé en deux parties, l'une périphérique
et l'autre profonde. Cette subdivision est purement physiologique,
car il n'existe entre les deux régions aucune limite morphologique.
La région périphérique contient les centres
germinatifs. La région profonde est un tissu lymphoïde
diffus...
Le cortex ganglionnaire
est habituellement partagé en deux parties, l'une périphérique
et l'autre profonde. Cette subdivision est purement physiologique,
car il n'existe entre les deux régions aucune limite morphologique.
La région périphérique contient les centres
germinatifs. La région profonde est un tissu lymphoïde
diffus...
L'importance relative des deux régions dans un ganglion
varie en fonction de son état d'activité et du type
de réaction immunitaire qui prédomine à un
moment donné.
Le centre germinatif
est une masse sphérique ou ovoïde qui en augmentant
de volume repousse et comprime les mailles du tissu réticulé.
Dans un centre actif, bien développé, on distingue
un pôle plus clair (1), toujours orienté vers
le sinus marginal, et un pôle plus sombre (2). Autour
du centre germinatif, il existe une couronne de petits lymphocytes
(3), très épaisse au pôle clair où
elle forme une espèce de "coiffe" et qui s'amenuise
à mesure que l'on se rapproche du pôle plus sombre.
Le centre germinatif et la couronne de petits lymphocytes qui
l'entourent sont encore décrits ensemble sous le nom de
nodule ou follicule lymphoïde secondaire. Les
cellules lymphoïdes du centre et de la couronne
appartiennent presque toutes à la lignée B;
l'ensemble de ces nodules lymphoïdes constitue donc la zone
bursodépendante du ganglion.
La coiffe (c) est
composée de petits lymphocytes qui seraient des cellules
à mémoire B. Cellesci sont produites par
prolifération clonale à partir des immunoblastes
B du centre germinatif proprement dit. La coiffe contient aussi
des cellules dendritiques qui sont cependant masquées
par l'abondance des petits lymphocytes et donc invisibles dans
cette préparation.
Les cellules dendritiques
sont aussi nombreuses dans la partie claire du centre germinatif.
Nous les reconnaissons dans cette imprégnation argentique
à leur aspect étoilé ou "arborescent"
caractéristique. Entre ces cellules, on trouve surtout
des petits lymphocytes, dont certains sont des lymphocytes
T probablement auxiliaires, de rares macrophages et quelques immunoblastes.
Les immunoblastes
et leurs précurseurs immédiats prédominent
dans la région sombre. Leur cytoplasme riche en ribosomes
est intensément coloré par la pyronine.
Les immunoblastes
ont l'aspect de grands lymphocytes et sont les précurseurs
des plasmocytes et des cellules à mémoire B. Leurs
mitoses sont fréquentes et justifient l'appellation de
''zone fertile" attribuée parfois à
la région sombre.
Cette région sombre contient aussi des macrophages
aisément reconnaissables à leur cytoplasme chargé
d'inclusions de phagocytose parmi lesquelles on distingue des
noyaux pycnotiques de lymphocytes.
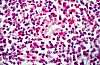 La partie interne du cortex,
sous les centres germinatifs, représente la zone thymodépendante
du ganglion. Cette zone s'insinue parfois entre les centres germinatifs
et peut même atteindre le sinus marginal. Elle est composée
de nombreux lymphocytes T localisés dans les mailles
du tissu réticulé. Elle doit son aspect diffus au
fait que les immunoblastes T restent dispersés parmi les
petits lymphocytes au lieu de se regrouper en nodules comme les
immunoblastes B. La zone thymodépendante contient encore
quelques macrophages et des cellules ''à interdigitations"
qui pour certains auteurs seraient les homologues des cellules
dendritiques des centres germinatifs. Elle est enfin caractérisée
par la présence des veinules postcapillaires.
La partie interne du cortex,
sous les centres germinatifs, représente la zone thymodépendante
du ganglion. Cette zone s'insinue parfois entre les centres germinatifs
et peut même atteindre le sinus marginal. Elle est composée
de nombreux lymphocytes T localisés dans les mailles
du tissu réticulé. Elle doit son aspect diffus au
fait que les immunoblastes T restent dispersés parmi les
petits lymphocytes au lieu de se regrouper en nodules comme les
immunoblastes B. La zone thymodépendante contient encore
quelques macrophages et des cellules ''à interdigitations"
qui pour certains auteurs seraient les homologues des cellules
dendritiques des centres germinatifs. Elle est enfin caractérisée
par la présence des veinules postcapillaires.
En dehors des zones lymphoïdes, le tissu réticulé
délimite les sinus du ganglion. Le sinus marginal
est une fente entre la capsule fibreuse et le cortex ganglionnaire.
Sa paroi externe est un épithélium pavimenteux
simple en continuité avec l'endothélium des
vaisseaux lymphatiques qui s'y déversent. Sa paroi interne
est une lame de cellules réticulées pavimenteuses,
percée de nombreux orifices qui permettent l'infiltration
des éléments de la lymphe dans le cortex. ganglionnaire.
La lumière du sinus est cloisonnée par de nombreuses
cellules réticulées attachées aux cellules
des parois.
Les sinus médullaires
sont délimités par des cellules réticulées
aux multiples prolongements qui agissent comme des déflecteurs
ralentissant considérablement le flux lymphatique. Ils
forment un réseau complexe de cavités tortueuses
qui contiennent des cellules libres, lymphocytes, macrophages
et granulocytes.
Dans cette préparation en microscopie à balayage,
la cellule qui se trouve dans le sinus est un macrophage.
On peut l'identifier parce que sa membrane est hérissée
de prolongements épais dont le centre est déprimé.
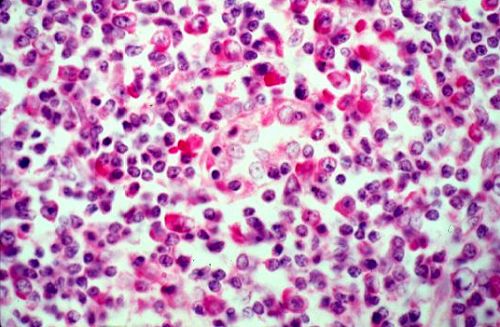
Dans une coupe histologique, les macrophages
se reconnaissent à leur noyau souvent excentrique, à
leur membrane irrégulière mais surtout aux inclusions
et vacuoles de leur cytoplasme. Les petites cellules à
noyau dense sont des lymphocytes.
Au niveau du hile ganglionnaire, les sinus médullaires
communiquent directement avec le début des voies lymphatiques
efférentes (V).
Autour des prolongements médullaires, les parois des sinus
forment une nappe percée d'orifices qui enveloppe étroitement
le tissu lymphoïde. La plupart des cellules situées
dans les prolongements appartiennent à la lignée
des plasmocytes.
Ces prolongements doivent donc être considérés
comme des régions mixtes de circulation cellulaire où
prédominent cependant les cellules de la lignée
B.
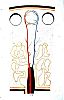 La vascularisation
d'un ganglion présente plusieurs particularités.
Le sang artériel lui est fourni par une artère
hilaire (1), qui se ramifie dans les travées conjonctives.
Les artérioles qui proviennent de ces ramifications
(2) pénètrent dans les prolongements médullaires
et fournissent un réseau capillaire diffus dans
tout le cortex et concentrique autour des centres germinatifs.
Quelques branches de ce réseau irriguent les centres germinatifs
eux-mêmes. Les capillaires sont drainés par de courts
segments veineux, les veinules postcapillaires situées
dans le cortex profond (3)...
La vascularisation
d'un ganglion présente plusieurs particularités.
Le sang artériel lui est fourni par une artère
hilaire (1), qui se ramifie dans les travées conjonctives.
Les artérioles qui proviennent de ces ramifications
(2) pénètrent dans les prolongements médullaires
et fournissent un réseau capillaire diffus dans
tout le cortex et concentrique autour des centres germinatifs.
Quelques branches de ce réseau irriguent les centres germinatifs
eux-mêmes. Les capillaires sont drainés par de courts
segments veineux, les veinules postcapillaires situées
dans le cortex profond (3)...
Cellesci deviennent ensuite les veines des prolongements
médullaires (4), puis les veines des septa conjonctifs
qui confluent vers le hile (5).
Tous ces vaisseaux, sauf les veinules postcapillaires, ont l'aspect
des vaisseaux d'autres organes. Voici une artériole
dans une travée conjonctive. On la reconnaît à
sa tunique vasculaire.
Les flèches indiquent la fin d'une travée conjonctive.
A cet endroit, l'artériole quitte la travée et pénètre
dans un prolongement médullaire. Plus loin, elle se ramifie.
La paroi de la veinule postcapillaire
est formée de cellules endothéliales larges,
presque cubiques et est infiltrée de lymphocytes. Cette
infiltration lymphocytaire a été diversement
interprétée. Pour la majorité des auteurs,
les lymphocytes quitteraient la circulation sanguine à
cet endroit électif. Pour d'autres au contraire, l'infiltration
lymphocytaire traduirait la pénétration des lymphocytes
ganglionnaires dans la lumière du vaisseau. Il est possible
que les deux processus coexistent mais il est vraisemblable que
dans ce cas, la récupération des lymphocytes à
partir du sang l'emporte. La veine postcapillaire représentée
ici a été observée dans une coupe épaisse
d'un 1/2 micron.
La voici telle qu'elle se présente dans une coupe classique
de 5 microns et colorée selon les techniques trichromiques
habituelles...
La lumière est étroite, l'endothélium possède
de larges cellules. Entre ces cellules et à leur base,
on observe les petits noyaux denses des lymphocytes qui
traversent la paroi vasculaire. Le vaisseau est souvent séparé
du tissu lymphoïde voisin par un fin liseré conjonctif.
Très rapidement, la veinule postcapillaire devient
une veinule normale. Voici cette transition.
A gauche, les cellules endothéliales sont épaisses
et font largement saillie dans la lumière du vaisseau.
Les noyaux denses des lymphocytes sont bien visibles dans la paroi.
A droite, la lumière vasculaire est limitée par
l'endothélium pavimenteux habituel.
L'infiltration lymphocytaire et l'épaisseur de l'épithélium
permettent de différencier la veinule postcapillaire des
autres veinules,
qu'elles se trouvent dans le tissu lymphoïde luimême
ou, comme celleci, dans un septum conjonctif. Leur endothélium
est aplati et leur paroi est dépourvue de fibres musculaires
lisses.
La circulation sanguine
intervient dans la recirculation des lymphocytes. En effet, la
lymphe contient beaucoup plus de lymphocytes en sortant d'un ganglion
qu'en y entrant. Ce supplément provient de deux sources.
La plus importante (95 %) est la circulation sanguine.
Les lymphocytes quitteraient le sang en traversant l'endothélium
des veines postcapillaires. Le reste provient de mitoses
au sein même du tissu lymphoïde.
La lymphe efférente d'un ganglion non stimulé contient,
quasi exclusivement, des lymphocytes T qui proviennent
de la région thymodépendante et forment le
groupe des lymphocytes ''recirculant". Les lymphocytes B
de la région bursodépendante sont moins mobiles
et stagnent dans le ganglion...
La fonction du ganglion est double. Il filtre la lymphe
qu'il reçoit et comme tout organe lymphoïde périphérique
produit les cellules immunitaires. Ces deux fonctions sont
très étroitement liées. La lymphe drainée
provient du liquide interstitiel.
Ceci peut être démontré en marquant un territoire
à l'aide d'un produit coloré
qui, injecté dans le derme par exemple, diffuse d'abord
dans les espaces du tissu conjonctif.
Après quelque temps, le produit coloré atteint et
colore les petits vaisseaux lymphatiques périphériques
qui sont les voies afférentes des ganglions les
plus proches. Les radiologues utilisent cet artifice pour repérer
les voies lymphatiques lorsqu'ils désirent réaliser
une lymphographie.
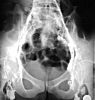
L'injection d'un produit de contraste dans ces lymphatiques périphériques
met en évidence les nombreuses voies afférentes
longues et grêles situées dans le bas de la radiographie,
les ganglions inguinaux euxmêmes et le vaisseau lymphatique
efférent beaucoup plus volumineux qui sort du hile ganglionnaire.
La lymphe amenée
par les lymphatiques afférents (1) est déversée
dans le sinus marginal (2). Elle y est déjà
considérablement ralentie. Une partie traverse les sinus
intermédiaires (3) et aboutit au labyrinthe des sinus
médullaires (4) où la complexité même
du trajet permet un brassage tel que chaque particule contenue
dans la lymphe entre en contact avec les macrophages de la région.
Une autre partie de la lymphe profuse à travers la paroi
du sinus marginal ou des sinus intermédiaires et diffuse
lentement à travers le cortex ganglionnaire avant d'aboutir
aux sinus médullaires. Ceuxci confluent avec le sinus
marginal du hile et forment à ce niveau un plexus de vaisseaux
tortueux qui pénètrent dans la capsule et sont drainés
par un vaisseau lymphatique efférent (5).
Le filtre ganglionnaire est efficace parce que les particules
(bactéries, substances étrangères, débris,
etc...) sont phagocytées par les macrophages. La
plupart d'entre elles sont dégradées mais certaines,
en particulier les substances minérales, ne le sont pas.
Ainsi les poussières absorbées lors de la respiration,
sont drainées par la lymphe pulmonaire vers les ganglions
thoraciques, bien qu'une part importante soit éliminée
par les expectorations. Les poussières qui arrivent aux
ganglions sont absorbées par les macrophages et y restent.
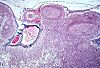
Si la quantité de poussières est importante, il
se forme un lacis anthracosique
le long de tous les sinus ganglionnaires, tel qu'on l'observe
dans cette préparation.
Un seul filtre est le plus souvent insuffisant. C'est pourquoi,
les ganglions sont disposés en chaînes. Une
lymphe particulièrement chargée d'impuretés
arrive totalement épurée au canal thoracique ou
à la grande veine lymphatique droite après être
passée par cette succession de filtres.
Dans des cas pathologiques, le filtre ganglionnaire est parfois
déficient. Dans certaines infections dont la tuberculose
les macrophages ganglionnaires ne peuvent tuer le bacille.
Ils s'associent, formant des cellules géantes
qui se nécrosent. Les zones de nécrose s'étendent
progressivement. Le ganglion perd sa capacité de filtre
et devient une source de dissémination bacillaire.
Dans d'autres circonstances, le ganglion ne réagit pas
aux éléments qu'il reçoit. Ici par exemple,
quelques cellules épithéliales cancéreuses
se sont insinuées dans le sinus marginal. Elles proliféreront
et métastaseront d'abord dans d'autres ganglions, puis
dans d'autres organes.
Le rôle épurateur du ganglion est à
l'origine de sa deuxième fonction. En effet, de nombreuses
substances filtrées sont antigéniques; elles sont
phagocytées par les macrophages. Une partie forme des complexes
antigèneanticorps qui sont retenus à la surface
des cellules dendritiques. Là, ils déclenchent une
stimulation lymphocytaire. Le ganglion, organe lymphoïde
périphérique, intervient dans l'immunité
humorale et dans l'immunité cellulaire. Selon le type de
réaction immunitaire, les modifications ganglionnaires
prédominent soit dans la zone bursodépendante, soit
dans la zone thymodépendante.
Une immunisation à médiation humorale peut
être obtenue par l'injection d'un antigène thymoindépendant,
par exemple un polysaccharide bactérien. Quelques heures
après l'injection de l'antigène, les ganglions qui
drainent le territoire où l'antigène est administré,
gonflent suite à une augmentation importante du flux
sanguin par vasodilatation...
Celleci est vraisemblablement déclenchée par
des médiateurs libérés au cours de
la réaction inflammatoire au lieu d'injection et qui atteignent
le ganglion avec la lymphe afférente.
A partir du deuxième jour, on observe une néoformation
vasculaire qui contribue à l'augmentation du flux sanguin.
Dès les premières heures, des granulocytes apparaissent
dans les sinus et infiltrent le tissu lymphoïde. Ils disparaissent
généralement après deux ou trois jours.
Au quatrième jour, se manifestent les premières
modifications au niveau des centres germinatifs. Ceuxci
augmentent de volume à la suite d'une prolifération
d'immunoblastes, induite par l'antigène. Ce développement
des centres germinatifs accroît encore la taille des ganglions
qui deviennent perceptibles au toucher et sont alors décrits
par les cliniciens sous le nom d'adénopathies. Au
début, les immunoblastes sont distribués de façon
uniforme dans les centres germinatifs. Progressivement, ils s'accumulent
au pôle basal, tandis que les cellules nouvellement formées,
à l'aspect de plasmocytes immatures, migrent vers le pôle
apical.
Les centres germinatifs prennent alors l'aspect typique décrit
plus haut, aspect qu'ils conservent pendant quelques semaines,
parfois même quelques mois, tant que dure la stimulation
antigénique. Celleci est entretenue par les complexes
antigèneanticorps fixés sur les prolongements
des cellules dendritiques.
Tout au long de la période d'immunisation, les précurseurs
plasmocytaires quittent les centres germinatifs, traversent le
cortex profond et s'accumulent comme ici dans les prolongements médullaires
pour y poursuivre leur différenciation en plasmocytes mûrs.
Bon nombre d'entre eux cependant quittent le ganglion par le vaisseau
lymphatique efférent avant d'être différenciés...
Ils aboutissent dans les prolongements médullaires d'autres
ganglions, dans d'autres organes et autour du foyer inflammatoire
où ils achèvent leur maturation. Dès le début
de leur différenciation dans le ganglion, ils produisent
des anticorps qui sont déversés dans la lymphe
efférente.
L'application locale d'oxazolone provoque une immunisation
à médiation exclusivement cellulaire. Au niveau
des ganglions de drainage, celleci se traduit par une turgescence
précoce de l'endothélium des veinules post-capillaires
et un recrutement accru de lymphocytes à ce niveau. Un
à deux jours après l'administration de l'antigène,
des immunoblastes groupés en îlots autour
de cellules à interdigitations apparaissent dans le cortex
profond. Les immunoblastes prolifèrent jusqu'au quatrième
jour environ et donnent naissance à une nouvelle population
de petits lymphocytes. Vers le septième jour, les immunoblastes
ont disparu, mais le cortex interne a considérablement
augmenté de volume. Les contacts intermembranaires
étroits observés entre cellules à interdigitations
et cellules T suggèrent l'intervention des cellules à
interdigitations dans la stimulation antigénique.
Les deux diapositives suivantes comparent des ganglions ayant
subi une stimulation antigénique de type humoral et de
type cellulaire.
Voici une stimulation de type humoral.
Le cortex est rempli de centres germinatifs. La médullaire
est réduite.
Voici une stimulation de type cellulaire.
Les centres germinatifs sont petits et rares. La zone interne
du cortex est considérablement élargie.
Cependant, une stimulation antigénique est rarement simple
et la plupart des réactions immunitaires se répercutent
à la fois dans les zones bursodépendantes et thymodépendantes.
L'exemple d'une stimulation antigénique complexe est celle
produite par le vaccin antituberculeux tué. Au cours des
premières heures qui suivent la vaccination, on observe
la présence dans les sinus ganglionnaires d'un nombre accru
de granulocytes, de lymphocytes et de macrophages. Au fil des
heures, le nombre des granulocytes diminue et celui des macrophages
augmente. A partir de la seizième heure, ceuxci se
regroupent en amas dans le sinus marginal et les sinus médullaires.
Les amas de macrophages deviennent plus compacts après
deux jours au point de bloquer certains sinus. Ils s'organisent
finalement en amas de cellules épithélioïdes
qui ne se désagrègent qu'après plusieurs
semaines. Les cellules épithélioïdes sont des
macrophages qui ont acquis des caractères morphologiques
de cellules sécrétrices, mais on ignore encore la
nature de leur sécrétion et leur rôle dans
l'immunité. Des stimulations antigéniques aussi
complexes s'accompagnent aussi d'une hyperplasie des centres germinatifs.
Nous avons terminé ainsi l'étude du ganglion lymphatique.
La leçon suivante est consacrée
à la rate.
 Les ganglions lymphatiques
sont de petits organes ovoïdes situés sur le
trajet des voies lymphatiques. Les voici tels que les radiologues
les observent après avoir injecté dans un vaisseau
lymphatique une substance opaque aux R.X. Ils sont parfois isolés,
mais le plus souvent groupés là où confluent
les lymphatiques d'un territoire. Les lymphatiques efférents
et les vaisseaux sanguins, abordent ou quittent le ganglion au
niveau du hile.
Les ganglions lymphatiques
sont de petits organes ovoïdes situés sur le
trajet des voies lymphatiques. Les voici tels que les radiologues
les observent après avoir injecté dans un vaisseau
lymphatique une substance opaque aux R.X. Ils sont parfois isolés,
mais le plus souvent groupés là où confluent
les lymphatiques d'un territoire. Les lymphatiques efférents
et les vaisseaux sanguins, abordent ou quittent le ganglion au
niveau du hile.