Les organes lymphoïdes (cours 1)
Le système lymphoïde est l'ensemble des cellules
capables d'identifier une molécule ou un organisme vivant
comme étranger (antigène) et de mettre en
oeuvre différents mécanismes capables de le détruire
ou d'inhiber son action.
Les cellules fondamentales du système lymphoïde sont
les lymphocytes, ils sont répartis dans l'organisme
de différentes façons. Ils peuvent être dispersés,
groupés dans des nodules lymphoïdes ou encore rassemblés
dans des organes particuliers.
D'autres types cellulaires coopèrent avec les lymphocytes
dans l'identification ou la destruction des antigènes.
Il existe deux types d'immunités. L'immunité
humorale dépend d'anticorps circulants produits à
distance de leur site d'action et donc aisément transmissibles
par le sérum. Selon le type de réaction des anticorps
avec leurs antigènes, on distingue différents types
d'anticorps : les anticorps cytotoxiques, tels que les hémolysines
dirigées contre des antigènes érythrocytaires,
les anticorps opsonisants qui permettent la phagocytose de certaines
bactéries, les anticorps précipitants et les anticorps
agglutinants tels que les hémagglutinines.
L'immunité cellulaire dépend d'anticorps
sessiles, sertis dans la membrane des cellules qui les produisent.
Dans certains cas, de très faibles quantités d'anticorps
sont malgré tout libérées mais, même
alors, cette immunité ne peut être transmise que
par des transferts cellulaires.
L'immunité cellulaire est responsable du rejet des greffes
et des tumeurs, de la réaction du greffon contre l'hôte,
des phénomènes d'hypersensibilité retardée
et des réactions vis-à-vis des virus et de certains
microorganismes intracellulaires.
Les organes lymphoïdes sont classés en organes
centraux, qui sont les sites de différenciation des
lymphocytes dans l'immunité cellulaire ou humorale. Le
thymus est l'organe lymphoïde central de l'immunité
cellulaire et la bourse de Fabricius est, chez les oiseaux,
l'organe central de l'immunité humorale.Son homologue chez
les mammifères est une portion de la moelle osseuse.
Les organes lymphoïdes périphériques
sont communs aux deux types d'immunités. Ce sont principalement
les ganglions, la rate et les structures lymphoïdes du tube
digestif. Ces organes sont partagés en aires thymo- et
bursodépendantes et les zones thymodépendantes rassemblent
les cellules responsables de l'immunité cellulaire ou cellules
T. Les zones bursodépendantes sont occupées
par les cellules effectrices de l'immunité humorale ou
cellules B.
Les lymphocytes sont produits dans la moelle, se différencient
dans les organes centraux, puis recirculent dans le sang et les
organes lymphoïdes périphériques. En cas de
stimulation par un antigène, certains lymphocytes se multiplient
très rapidement, hors de la moelle osseuse.
Le thymus est un organe lymphoïde central de l'immunité
cellulaire. En effet, l'organe lymphoïde central de l'immunité
humorale n'est connu que chez les oiseaux. Chez eux, il est une
petite poche lympho-épithéliale annexée au
cloaque, la bourse de Fabricius.
 Le thymus est situé
dans le médiastin antérieur et supérieur.
Il est composé de deux lobes pyramidaux dont la base repose
sur le péricarde et dont le sommet se prolonge dans la
partie inférieure du cou. Chez le nouveau-né, il
mesure environ 5 cm de long, 3 cm de large et 1 cm en épaisseur
et pèse environ 10 grammes. Sa consistance est molle et
sa couleur, grisâtre chez l'enfant, devient jaunâtre
chez l'adulte parce que l'organe est progressivement infiltré
de tissu adipeux.
Le thymus est situé
dans le médiastin antérieur et supérieur.
Il est composé de deux lobes pyramidaux dont la base repose
sur le péricarde et dont le sommet se prolonge dans la
partie inférieure du cou. Chez le nouveau-né, il
mesure environ 5 cm de long, 3 cm de large et 1 cm en épaisseur
et pèse environ 10 grammes. Sa consistance est molle et
sa couleur, grisâtre chez l'enfant, devient jaunâtre
chez l'adulte parce que l'organe est progressivement infiltré
de tissu adipeux.
Chaque lobe est enveloppé d'une capsule fibreuse.
De la face interne de cette capsule partent des septas conjonctifs
qui s'enfoncent plus ou moins profondément dans le parenchyme
et le divisent en nombreux lobules. L'image montre plusieurs lobules
dans un fragment de lobe.
Chaque lobule comprend une zone périphérique sombre,
le cortex, et une région centrale claire, la médullaire
dans laquelle on distingue de petites zones colorées en
rouge, les corpuscules de Hassall. Les septas conjonctifs
interlobulaires n'isolent que la région corticale, si bien
que la médullaire d'un lobule se prolonge dans la médullaire
de ses voisins.
A plus fort grossissement,
l'aspect sombre du cortex des lobules thymiques est bien visible,
il est dû à sa richesse particulière en petites
cellules à noyau dense. Le lobule peut être considéré
comme l'unité morphologique et fonctionnelle du thymus.
Sa fonction est de produire les lymphocytes T, cellules
effectrices de l'immunité cellulaire.
La trame du lobule
est un réseau de cellules épithéliales
étoilées, arrimées les unes aux autres
au niveau de leurs prolongements. Les mailles de ce réseau
épithélial sont larges dans le cortex et plus étroites
dans la médullaire. En périphérie, le réseau
est achevé par une assise continue de cellules aplaties
séparées du tissu conjonctif de la capsule, des
septas et des espaces périvasculaires par une membrane
basale. Les mailles corticales contiennent surtout des
petits lymphocytes, appelés thymocytes, et quelques
macrophages. Les mailles médullaires abritent une
population beaucoup plus polymorphe, comprenant des lymphocytes,
des cellules épithéliales globuleuses, des macrophages
et quelques rares granulocytes. En résumé, le thymus
est comparable à une éponge épithéliale
remplie principalement de lymphocytes.
Dans une coupe histologique classique,
les cellules épithéliales de la charpente thymique,
marquées d'une flèche dans cette préparation,
se reconnaissent grâce à leur noyau volumineux
et pâle qui contraste avec les petits noyaux denses
des lymphocytes. L'organisation en réseau n'y apparaît
pas, car les prolongements cytoplasmiques, longs et grêles,
des cellules épithéliales sont masqués par
l'abondance des cellules lymphoïdes. On constate cependant
que la composante épithéliale est plus importante
dans la médullaire, à gauche dans l'image, que dans
le cortex.
Bien qu'en continuité l'un avec l'autre, le réseau
cortical et le réseau médullaire sont constitués
de cellules étoilées différentes. Les cellules étoilées corticales
possèdent un noyau volumineux et pâle. La chromatine
y existe principalement sous forme d'euchromatine. L'hétérochromatine
n'est représentée que par une mince bande et quelques
blocs denses accolés à l'enveloppe nucléaire.
Le gros nucléole spongieux est formé par un nucléolonème
épais. Le cytoplasme contient des vésicules caractéristiques
dont la taille est comparable à celle des mitochondries.
Elles sont entourées d'une membrane simple. Certaines paraissent
vides, d'autres contiennent un matériel floconneux dense
aux électrons. Le nombre de ces vésicules diminue
à l'âge adulte. Le réticulum endoplasmique
rugueux et l'appareil de Golgi sont relativement peu développés.
Toutes les cellules possèdent des faisceaux de microfilaments
et sont attachées les unes aux autres par des desmosomes.
Les cellules étoilées de la médullaire
ressemblent aux cellules corticales par leur forme, leur disposition
en réseau, leurs microfilaments et leurs desmosomes. Elles
en diffèrent cependant à certains points de vue.
Les cellules sont plus petites. Le noyau est plus allongé,
les blocs d'hétérochromatine y sont plus grands
et plus abondants, le nucléole est plus compact. Les vésicules
cytoplasmiques sont très rares. Les cellules étoilées
médullaires sont en rapport étroit avec un autre
type de cellules épithéliales, les cellules globuleuses,
qu'elles enveloppent de leurs prolongements et auxquelles elles
sont attachées par des desmosomes.
Les cellules étoilées médullaires
s'associent parfois en couches concentriques, formant alors
des mailles plus serrées. Le nombre des interdigitations
et desmosomes augmente fortement. Les cellules sont très
riches en microfilaments et contiennent souvent des grains de
kératohyaline. Au centre de ces associations se trouvent
des débris soit de lymphocytes, soit de cellules globuleuses,
soit encore de cellules inflammatoires, neutrophiles, éosinophiles
ou macrophages. Chacune de ces associations représente
un corpuscule de Hassall, caractéristiques de la
médullaire et aisément reconnaissables parce que
leurs lamelles cellulaires épithéliales sont disposées
comme les écailles dans un bulbe d'oignon. Leur importance
varie cependant d'une espèce à l'autre : ils sont
réduits chez le rat et la souris et fort développés
chez l'homme, le cobaye et le chien. Aucune fonction ne leur est
reconnue.
D'autres cellules épithéliales, de forme généralement
arrondie, se trouvent dans les mailles du réseau médullaire.
Les cellules épithéliales globuleuses
possèdent un grand noyau pâle, identique à
celui des cellules étoilées corticales. Mais leurs
vésicules cytoplasmiques ont une localisation, une structure
et un contenu différents. Elles sont nombreuses et groupées
dans la région juxtanucléaire, repoussant le noyau
en position excentrique. Elles contiennent un matériel
amorphe de densité variable, à moins que celui-ci
n'ait été extrait lors de la préparation
du tissu. Elles sont limitées par une membrane simple dont
naissent des microvillosités et parfois des cils qui se
projettent dans la lumière.
Les vésicules des cellules globuleuses
peuvent fusionner et former ainsi de véritables microkystes
ciliés intracellulaires.
Des cellules voisines peuvent s'associer et, en fusionnant leurs
vésicules, donner naissance à des kystes "extracellulaires".
Dans le réseau et parmi les cellules globuleuses, certaines cellules
sont plurinucléées. Leur nature épithéliale
est confirmée par la présence de microfilaments
et de desmosomes. Leur cytoplasme est pauvre en vésicules.
Les cellules sont peu fréquentes à la naissance,
mais augmentent lorsque le thymus involue ou en cas d'affection
auto-immunitaire.
La fonction de l'épithélium thymique est double.
D'une part, les cellules étoilées du réseau
ont un rôle de soutien car elles forment la charpente
permanente de l'organe. D'autre part, les cellules étoilées
corticales et les cellules globuleuses médullaires assurent
le microenvironnement nécessaire à la multiplication,
à la différenciation et à l'acquisition de
la compétence immunitaire des lymphocytes T. Les cellules
étoilées corticales entourent de leur prolongements
les lymphocytes créant ce microenvironnement, on les appelle
souvent des cellules "nurses". On ne connaît
pas le mécanisme exact par lequel les cellules épithéliales
s'acquittent de ces tâches, mais il est certain qu'il implique
l'action de cytokines sécrétées localement
et d'hormones. Quatre facteurs d'origine thymique ont déjà
été isolés : le facteur sérique, la
thymosine, la thymopoïétine et le facteur thymique
humoral. Tous sont des substances peptidiques dont l'administration
à des animaux privés de thymus dès la naissance
produit des effets superposables à ceux résultant
de la greffe de l'organe entier.
Les lymphocytes thymiques
dérivent de lymphoblastes originaires de la moelle osseuse.
Les lymphoblastes, qui constituent la population lymphoïde
"pré-thymique", gagnent le thymus par voie sanguine
et s'y accumulent dans la région sous-capsulaire du
cortex où ils se divisent fréquemment. Les lymphoblastes
ont l'aspect de grands lymphocytes. Leur accumulation dans la
région sous-capsulaire rend cette zone moins colorable,
dans cette préparation colorée au bleu de toluidine.
L'intensité de la multiplication des lymphoblastes périphériques
peut être évaluée par la fréquence
d'incorporation d'un précurseur radioactif de l'ADN, la
thymidine tritiée. L'autoradiographie
que voici montre que cette substance est incorporée dans
la zone sous-corticale plus que partout ailleurs.
Les divisions successives
des lymphoblastes donnent naissance aux petits thymocytes à
noyau dense, qui, tout en se divisant à un rythme moindre,
migrent dans la région profonde du cortex (1). Certains
dégénèrent et sont phagocytés par
les macrophages. On admet généralement que les autres,
après avoir cessé leurs divisions, migrent dans
la médullaire thymique puis quittent le thymus en empruntant
les veinules de la jonction cortico-médullaire pour coloniser
les aires thymodépendantes des organes lymphoïdes
périphériques: manchons périartériolaires
de la rate et zones corticales particulières des ganglions
lymphatiques. L'autre théorie (2) proposant que quelques
lymphoblastes issus de la moelle gagnent la zone médullaire
sans passer par le cortex, s'y divisent et s'y différencient
en une population de lymphocytes différente de la population
corticale et destinée aux organes périphériques
n'a pu être démontrée.
Les lymphocytes corticaux et médullaires sont différents.
Les lymphocytes corticaux n'ont pas de compétence
immunitaire, sont détruits par l'hydrocortisone et le rythme
de leurs divisions est rapide. Leur noyau est rond et riche en
hétérochromatine. Leur cytoplasme peu abondant est
riche en ribosomes mais pauvre en autres organites. Au cours de
leur différenciation, l'activation de leur génome
induit l'apparition d'antigènes membranaires spécifiques
des cellules T mais ils ne possèdent pas les antigènes
d'histocompatibilité.
Les lymphocytes médullaires ont la compétence
immunitaire, sont insensibles à l'hydrocortisone et se
divisent rarement. Leur noyau est échancré, leur
cytoplasme, plus abondant, contient un système de Golgi
bien développé et de nombreux ribosomes. Le motif
antigénique de leur membrane est modifié par rapport
à celui des thymocytes corticaux; certains antigènes
régressent d'autres, comme les antigènes d'histocompatibilité,
augmentent.
Les lymphocytes médullaires ne possèdent pas toutes
les propriétés antigéniques des lymphocytes
T circulants. La population "post-thymique" achève
sa différenciation, sous l'influence de facteurs thymiques,
dans les organes lymphoïdes périphériques.
Cette population "post-thymique" existe déjà
dès la naissance et est entretenue par le passage continu
de nombreux lymphoblastes dans le thymus.
Les macrophages
sont très nombreux. Leur aspect dépend essentiellement
de leur état fonctionnel et de leur contenu. Ils interviennent
dans l'élimination des cellules mortes. Leur cytoplasme
peut donc contenir des thymocytes, des cellules épithéliales
ou les produits de leur dégradation. En microscopie optique,
ils ressemblent parfois aux cellules épithéliales
mais l'absence de desmosomes et de microfilaments permet de les
reconnaître alors aisément en microscopie électronique.
Les macrophages sont particulièrement concentrés
autour des vaisseaux thymiques.
La circulation sanguine
est assurée par les artères interlobulaires
(a) qui cheminent dans les septa conjonctifs des lobes thymiques.
Les artères proviennent des artères mammaires internes
et des artères thyroïdiennes inférieures. Elles
donnent naissance aux artérioles intralobulaires
(b) situées à la frontière corticomédullaire.
Cellesci se ramifient en capillaires qui s'enfoncent dans
le cortex thymique et forment des arcades périphériques
anastomosées en un réseau cortical (c). Quelques
capillaires quittent le cortex et sont drainés par les
veines de la capsule conjonctive. Un petit nombre de capillaires
irrigue la médullaire (d). La majorité des
capillaires sont récurrents et se dirigent vers la jonction
corticomédullaire où ils confluent en veinules
postcapillaires (e) qui se réunissent en veines
intramédullaires (f) pour quitter l'organe par les
septas conjonctifs.
Voici la coupe d'un thymus
où la différence entre la vascularisation corticale
et la vascularisation médullaire est accentuée parce
que l'organe a été fixé par perfusion. L'importance
des capillaires dans la zone corticale et des veinules
dans la médullaire est clairement démontrée
Comme dans les autres organes lymphoïdes, la veinule postcapillaire
se reconnaît non seulement à son épithélium
cubique très différent de l'endothélium
des autres veinules mais aussi à la présence des
lymphocytes qui s'insinuent entre deux cellules épithéliales
adjacentes et pénètrent ainsi dans la circulation
sanguine.
Les thymocytes en voie de différenciation sont protégés
de tout contact avec une substance étrangère par
la barrière hématothymique.
Celleci est purement corticale. Elle est composée
d'une double paroi. La paroi interne est l'endothélium
des capillaires et des veinules postcapillaires imperméable
à tout traceur. Les cellules endothéliales sont
unies les unes aux autres par des desmosomes et des jonctions
serrées qui assurent une imperméabilité presque
totale, sauf dans la région souscapsulaire où
les capillaires sont fenestrés. La paroi externe
est une couche continue de cellules épithéliales.
Au niveau des capillaires, les membranes basales des deux couches
sont soudées; au niveau des veinules postcapillaires,
elles sont séparées par des fibres conjonctives
et des péricytes. Les macrophages amassés autour
des vaisseaux renforcent le contrôle exercé par la
barrière hématothymique.
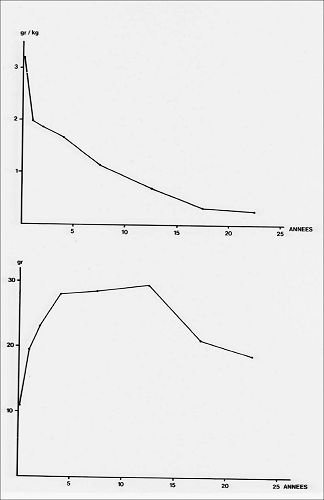
 Dans l'espèce humaine, le poids relatif
du thymus est maximal à la naissance puis diminue constamment.
Son poids absolu, au contraire, augmente au cours de l'enfance
et est maximal entre la onzième et la quinzième
année. Il décroît ensuite rapidement au cours
de l'adolescence puis lentement au cours de la vie adulte.
Dans l'espèce humaine, le poids relatif
du thymus est maximal à la naissance puis diminue constamment.
Son poids absolu, au contraire, augmente au cours de l'enfance
et est maximal entre la onzième et la quinzième
année. Il décroît ensuite rapidement au cours
de l'adolescence puis lentement au cours de la vie adulte.
Lors de l'involution, le parenchyme diminue mais le collagène
et le tissu adipeux interlobulaire augmentent. L'involution modifie
les proportions des différents constituants car elle atteint
préférentiellement le cortex.
Elle semble due avant tout à une diminution progressive
de la production des thymocytes. La trame épithéliale
est d'abord contractée par le départ des thymocytes
puis involue à son tour. Chez le vieillard, il ne reste
que des vestiges thymiques, surtout médullaires, inclus
dans un tissu graisseux. Dans cette image,
on peut comparer l'aspect du thymus involué à droite
à celui du thymus d'un enfant de deux ans.
De nombreux agents tels que la dénutrition, les infections,
l'irradiation induisent une involution aiguë mais réversible
du thymus. Elle est due à la disparition des lymphocytes
corticaux qui deviennent pycnotiques et sont phagocytés
par des macrophages. Dès que cesse la cause de cette involution
accidentelle, le thymus régénère en quelques
jours grâce aux lymphoblastes qui pénètrent
dans le cortex. L'involution accidentelle
implique l'intervention des hormones corticosurrénaliennes.
Ici, elle est expérimentale et provoquée par l'injection
d'hydrocortisone. Les lymphocytes médullaires persistent
parce qu'ils sont résistants à cette hormone. Ceci
explique l'image inversée : la médullaire est plus
sombre que la corticale parce que plus riche en lymphocytes.
La prochaine leçon sera consacrée
au ganglion lymphatique.
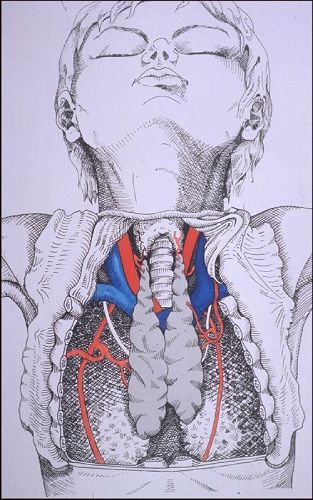 Le thymus est situé
dans le médiastin antérieur et supérieur.
Il est composé de deux lobes pyramidaux dont la base repose
sur le péricarde et dont le sommet se prolonge dans la
partie inférieure du cou. Chez le nouveau-né, il
mesure environ 5 cm de long, 3 cm de large et 1 cm en épaisseur
et pèse environ 10 grammes. Sa consistance est molle et
sa couleur, grisâtre chez l'enfant, devient jaunâtre
chez l'adulte parce que l'organe est progressivement infiltré
de tissu adipeux.
Le thymus est situé
dans le médiastin antérieur et supérieur.
Il est composé de deux lobes pyramidaux dont la base repose
sur le péricarde et dont le sommet se prolonge dans la
partie inférieure du cou. Chez le nouveau-né, il
mesure environ 5 cm de long, 3 cm de large et 1 cm en épaisseur
et pèse environ 10 grammes. Sa consistance est molle et
sa couleur, grisâtre chez l'enfant, devient jaunâtre
chez l'adulte parce que l'organe est progressivement infiltré
de tissu adipeux.