Les glandes endocrines (cours 3)
Après avoir étudié les principaux organes
endocrines, nous abordons un domaine plus complexe et en pleine
expansion: les cellules endocrines du système digestif.
Certaines cellules endocrines du pancréas et celles du
tube digestif présentent des particularités communes
et sont généralement regroupées dans un système
plus général, parfois dénommé système
APUD, qui comprend également des cellules endocrines
en dehors du tube digestif.
 Les îlots de Langerhans
sont répartis dans la masse exocrine du pancréas.
En coupe, ce sont des plages claires, rondes ou ovales constitués
d'un lacis cellulaire anastomosé et d'un riche réseau
de capillaires. Le diamètre des îlots est compris
entre 30 et 300 micromètres. Leur nombre, très élevé,
peut varier chez l'homme entre 200.000 et 2.300.000 .
Les îlots de Langerhans
sont répartis dans la masse exocrine du pancréas.
En coupe, ce sont des plages claires, rondes ou ovales constitués
d'un lacis cellulaire anastomosé et d'un riche réseau
de capillaires. Le diamètre des îlots est compris
entre 30 et 300 micromètres. Leur nombre, très élevé,
peut varier chez l'homme entre 200.000 et 2.300.000 .
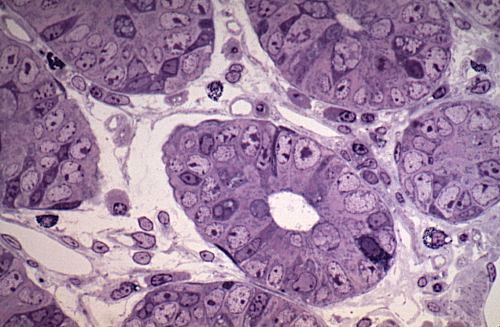
Voici un îlot de Langerhans,
à plus fort grossissement, dans une coupe colorée
selon la méthode trichromique classique. Les cellules présentent
les caractéristiques des cellules endocrines à
sécrétion protéique. Leur noyau central
est riche en euchromatine et le nucléole est bien apparent.
Leur cytoplasme à ce grossissement paraît homogène,
parce que les grains sont très petits. Les cellules sont
en relation étroite avec des capillaires, dont on voit
la lumière et l'endothélium.
Certaines colorations, notamment celle de Gomori,
permettent de distinguer deux types cellulaires: les cellules
A, sécrétrices de glucagon qui représentent
15 à 20% de la population cellulaire et les cellules
B, sécrétrices d'insuline qui représentent
60 à 80 % de la population cellulaire. Les granules des
cellules A ont une affinité élective pour les colorants
acides et sont colorés en rouge. Les granules
des cellules B sont sélectivement colorés en bleu.
En microscopie électronique,
les cellules A et B se reconnaissent uniquement à l'aspect
de leurs grains. Les grains des cellules A sont très
denses et leur diamètre varie entre 190 et 310 nm. Les
grains des cellules B ont un diamètre variant entre 225
et 375 nm. Ils contiennent une masse centrale très dense
aux électrons, qui est séparée de la membrane
par un large espace clair et leur aspect varie selon les espèces.
Ils peuvent être homogènes, de densité moyenne,
ou même contenir un ou plusieurs cristaux denses. Hormis
l'aspect des granules, les cellules A et B ont 1'ultrastructure
typique des cellules endocrines à sécrétion
protéique: large noyau riche en euchromatine, nucléole
bien apparent, complexe de Golgi développé.
Les grains des cellules A contiennent le glucagon
et sa préhormone, la glicentine; les grains des
cellules B contiennent de l'insuline. Cependant
l'insuline n'est pas stockée telle quelle; elle forme un
complexe avec le zinc, ce qui diminue sa solubilité
et explique la formation occasionnelle de cristaux. Le rôle
du zinc est d'assurer une plus grande stabilité aux complexes.
Outre les cellules A et B, le pancréas endocrine comprend
également des cellules D,
qui sécrètent la somatostatine. Voici leur
aspect en microscopie électronique. Les granules sont ronds
et leur densité aux électrons est plus faible que
celle des granules A et B.
Les cellules endocrines du pancréas sont en relation étroite
avec des capillaires fenestrés.
Cette cellule endocrine n'est séparée du capillaire
que par une double membrane basale, l'une épithéliale,
l'autre endothéliale. Les fenestrations des capillaires
sont bien visibles en coupe transversale.
La répartition des cellules
A, B et D au sein de l'îlot peut varier d'une espèce
à l'autre. Dans beaucoup d'espèces, dont l'homme,
les cellules A et D forment une couronne périphérique
autour des cellules B. La coexistence de ces trois types cellulaires
dans les îlots laisse supposer qu'il existe entre eux une
relation fonctionnelle. Il est, en effet, prouvé que la
somatostatine inhibe la sécrétion d'insuline
et de glucagon.
Outre les cellules A, B et D, six autres types cellulaires ont
été décrits dans le pancréas endocrine.
Ce sont les cellules PP, qui produisent le polypeptide pancréatique,
les cellules S qui produisent la sécrétine, les
cellules G sécrétant la gastrine, les cellules P
sécrétant la bombésine, les cellules EC qui
contiendraient des amines biogènes et les cellules Dl,
dont le produit est inconnu. Ces cellules ont été
identifiées principalement par des techniques d'immunofluorescence,
mais la relation entre l'ultrastructure et la nature du produit
de sécrétion n'est bien établie que pour
les cellules A, B, D et PP.
La sécrétion du glucagon
est stimulée par toute baisse de la glycémie,
qu'elle soit provoquée par le jeûne ou par
une décharge d'hydrates de carbone.
Le glucagon agit principalement sur les cellules hépatiques.
Sa liaison à leurs récepteurs active l'adénylcyclase
et augmente donc la formation d'AMP cyclique. Le glucagon stimule
la glycogénolyse et la gluconéogenèse
dans le foie et provoque ainsi une hausse de la glycémie.
L'insuline agit principalement sur les hépatocytes,
les adipocytes et les cellules musculaires. Elle
a des effets rapides et des effets tardifs.
Elle stimule rapidement le transport membranaire du glucose
et des acides aminés dans la cellule, en se liant à
des récepteurs membranaires spécifiques. L'effet
majeur de cette liaison est de faciliter la pénétration
du glucose. En soustrayant le glucose du sang, elle se comporte
donc comme une hormone hypoglycémiante, et en facilitant
la pénétration de glucose dans le foie, elle stimule
indirectement la synthèse de glycogène.
Plus tardivement, l'insuline active la synthèse d'ADN,
d'ARN et de protéines. Ces effets sont déclenchés
par la liaison de l'insuline à son récepteur, qui
est une tyrosine kinase qui phosphoryle diverses protéines.
La sécrétion d'insuline
est stimulée par toute élévation de la
glycémie, mais on ignore comment. Il est possible qu'un
métabolite du glucose ou un acide aminé induise
une accumulation intracellulaire de calcium ionisé, qui
en agissant sur les microtubules, stimulerait l'exocytose et la
sécrétion d'insuline. Le rôle des microtubules
est évident lorsqu'on étudie la sécrétion
d'insuline apres une stimulation par le glucose. En effet, cette
sécrétion est biphasique. Elle augmente rapidement
endéans les deux minutes qui suivent l'administration de
glucose, puis diminue. Une seconde augmentation survient après
huit minutes et atteint un plateau après cinquante minutes.
L'administration de vinblastine, inhibiteur des microtubules,
empêche la seconde phase de la sécrétion.
Donc, lors de la phase rapide, des grains de sécrétion
situés à proximité des membranes, libèreraient
rapidement leur insuline. Par contre, lors de la deuxième
phase, des grains de sécrétion devraient atteindre
les membranes et leur transport nécessiterait l'intervention
de microtubules.
En résumé, les cellules A et B du pancréas
ont des fonctions opposées. Les cellules A sécrètent
le glucagon dont les effets sont hyperglycémiants et les
cellules B sécrètent l'insuline dont les effets
sont hypoglycémiants. En fait, les cellules A et B fonctionnent
de manière synergique pour maintenir la glycémie
constante.
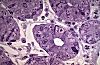 Les cellules endocrines isolées du tube digestif
sont dispersées dans la muqueuse depuis le cardia jusqu'au
rectum. Elles sont plus nombreuses dans les régions pylorique
et duodénale. Voici une cellule endocrine dans une coupe
de 1 micromètre, colorée au bleu de toluidine. Elle
est insérée entre les cellules exocrines d'une glande
de Lieberkuhn et se distingue de ses voisines par la position
des grains de sécrétion. Ceux-ci, très
colorés sont situés surtout au pôle
orienté vers les capillaires du chorion.
Les cellules endocrines isolées du tube digestif
sont dispersées dans la muqueuse depuis le cardia jusqu'au
rectum. Elles sont plus nombreuses dans les régions pylorique
et duodénale. Voici une cellule endocrine dans une coupe
de 1 micromètre, colorée au bleu de toluidine. Elle
est insérée entre les cellules exocrines d'une glande
de Lieberkuhn et se distingue de ses voisines par la position
des grains de sécrétion. Ceux-ci, très
colorés sont situés surtout au pôle
orienté vers les capillaires du chorion.
Certaines colorations spécifiques permettent de distinguer
ces cellules entre elles. Ce sont essentiellement les colorations
à l'argent, parmi lesquelles on distingue les réactions
argentaffines et les réactions argyrophiles.
La réaction argentaffine ne se produit que dans
les cellules endocrines contenant de la sérotonine;
elles réduisent directement l'argent ammoniacal
en argent métallique. Toutes les autres cellules endocrines
de la muqueuse digestive sont argyrophiles; elles fixent
les ions argent dont la réduction n'est possible qu'après
addition d'agents réducteurs. Les cellules endocrines
du pancréas, hormis les cellules B, le sont également.
Voici l'aspect de cellules argyrophiles
dont le pôle basal est coloré en noir.
On peut également différencier les cellules endocrines
isolées en fonction de leur rapport avec le milieu extérieur.
Dans le type fermé, la cellule n'est pas en contact
avec la lumière intestinale. Elle est enchâssée
entre les cellules exocrines, du côté basal de l'épithélium
digestif. Dans le type ouvert, la cellule est en rapport
avec la lumière. Elle est unie aux cellules exocrines par
des systèmes de jonctions.
Voici une cellule de type fermé.
Elle est piriforme. Son noyau rond est central. Les grains de
sécrétion de forme et de taille variables, sont
très denses et tous groupés du côté
du chorion. Les autres organites sont situés du côté
opposé. Ce type de cellule se retrouve surtout dans le
fond de l'estomac.
Voici une cellule de type ouvert.
Sa membrane apicale est garnie de microvillosités
en contact avec la lumière. La plupart des grains de sécrétion
sont rassemblés du côté du chorion. Ils sont
plus petits et leur forme est plus régulière que
dans le cas précédent, leur contenu est dense. Les
autres organites sont situés du côté de la
lumière.
Indépendamment de leur aspect morphologique, les cellules
endocrines du tube digestif peuvent encore être groupées
selon leur fonction. En effet, certaines sont purement
endocrines; leurs hormones gagnent directement la circulation
sanguine et agissent sur des organes cibles situés à
distance. D'autres, par contre, sont paracrines; leurs
hormones exercent une action purement locale, sans être
véhiculées par le sang. Certaines cellules sont
à la fois endocrines et paracrines.
La cellule I est
l'exemple d'une cellule purement endocrine. Elle est située
principalement dans l'intestin grêle et sécrète
la cholécystokinine qui est libérée
dans le sang et stimule la contraction de la vésicule biliaire.
La cellule I est de type ouvert.
L'exemple le mieux connu de cellule paracrine est la cellule D,
de type fermé dans le pancréas et le fond de l'estomac,
et de type ouvert dans l'intestin. Elle sécrète
la somatostatine qui n'entre pas dans la circulation sanguine
mais agit directement sur des cellules voisines. Dans le tube
digestif, elle inhibe notamment les sécrétions endocrines
de gastrine, d'entéroglucagon et de cholécystokinine.
Les cellules endocrines du tube digestif sont stimulées de manière différente
selon leur type.
Dans le type fermé, la stimulation est soit humorale,
soit nerveuse. Par contre, les cellules de type ouvert,
de loin les plus nombreuses surtout dans l'intestin, sont stimulées
par des substances de la lumière intestinale. Celles-ci
se fixent sur les microvillosités ou pénètrent
dans la cellule via des vésicules de pinocytose.
Les cellules du type ouvert
sont donc sensibles à toute variation du contenu de
la lumière intestinale. Les cellules S du bulbe duodénal,
par exemple, détectent rapidement une acidification du
contenu intestinal et libèrent alors de la sécrétine
qui stimule la production de substances alcalines par les glandes
intestinales, le foie et le pancréas. L'alcalinisation,
par l'ingestion de viande ou de bicarbonate stimule la production
de gastrine qui augmente la sécrétion d'acide par
l'estomac.
D'autres cellules de type ouvert sont sensibles à une augmentation
du contenu intestinal en glucose ou en acides gras. Elles sécrètent
alors un peptide appelé GIP ou "gastric inhibitory
peptide" qui inhibe la sécrétion de suc gastrique
et stimule la libération d'insuline.
Puisque les cellules de type ouvert sont à la fois sensorielles
et sécrétrices, certains les considèrent
comme des paraneurones. En effet, elles ressemblent à
des neurones car, comme eux, elles possèdent une zone sensorielle
spécialisée. Elles s'en différencient
par l'absence de connexion synaptique, leur produit étant
libéré dans le milieu intérieur.
Si certaines hormones produites dans la paroi du tube digestif
influencent la sécrétion de ses cellules exocrines,
d'autres ont comme fonction de régler sa motilité.
Toute perturbation dans leur sécrétion s'accompagne
d'un trouble. Ainsi l'hypersécrétion du facteur
appelé "vasoactive intestinal peptide" ou VIP
induit un relâchement des muscles lisses et une accélération
du transit intestinal, se traduisant par de la diarrhée.
En résumé, les cellules endocrines du tube digestif
peuvent présenter deux aspects morphologiques: elles peuvent
être de type fermé ou de type ouvert
. Elles sont caractérisées par leurs nombreux grains
de sécrétion, de taille, forme et densité
variables. Leur sécrétion peut être endocrine
ou paracrine et la plupart semblent à la fois sensorielles
et sécrétrices. Avec le système nerveux,
elles règlent la digestion.
L'endocrinologie du tube digestif est cependant plus compliquée
que ne le laisse supposer ce bref aperçu. En effet, depuis
ces dix dernières années, les découvertes
se succèdent à un rythme rapide. A présent,
de multiples hormones ou parahormones ont déjà été
extraites et purifiées; les cellules qui les produisent
n'ont été identifiées que dans certains cas.
Les cellules endocrines du tube digestif sont parfois considérées
comme faisant partie d'un système plus général
dénommé APUD. Ce terme est formé des
initiales des mots anglais "Amine content or Precursor Uptake
and Decarboxylation" qui définissent les propriétés
des cellules de ce système. Celles-ci peuvent, en effet,
contenir ou capter des précurseurs d'amines et les décarboxyler.
Ces précurseurs sont généralement la dihydroxyphénylalanine
ou le 5-hydroxytryptophane; la dopamine, la sérotonine
et l'histamine sont les trois amines qui ont pu être détectées.
Ces amines sont stockées dans les mêmes grains de
sécrétion que les hormones peptidiques.
Les cellules APUD ont toutes une même origine embryologique.
Elles dérivent des crêtes neurales et colonisent
le tube digestif et de nombreux autres organes comme la thyroïde,
le poumon, la trachée, les canaux biliaires, la prostate,
et l'urèthre.
Ceci termine l'étude des cellules glandulaires endocrines
du tube digestif . Au cours de la prochaine leçon,
nous terminerons notre analyse du système endocrinien.
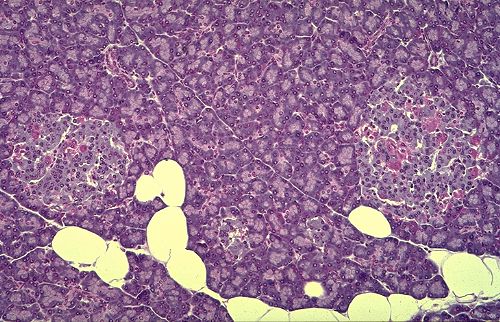 Les îlots de Langerhans
sont répartis dans la masse exocrine du pancréas.
En coupe, ce sont des plages claires, rondes ou ovales constitués
d'un lacis cellulaire anastomosé et d'un riche réseau
de capillaires. Le diamètre des îlots est compris
entre 30 et 300 micromètres. Leur nombre, très élevé,
peut varier chez l'homme entre 200.000 et 2.300.000 .
Les îlots de Langerhans
sont répartis dans la masse exocrine du pancréas.
En coupe, ce sont des plages claires, rondes ou ovales constitués
d'un lacis cellulaire anastomosé et d'un riche réseau
de capillaires. Le diamètre des îlots est compris
entre 30 et 300 micromètres. Leur nombre, très élevé,
peut varier chez l'homme entre 200.000 et 2.300.000 .