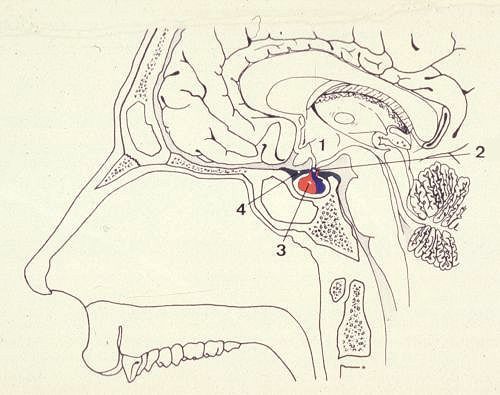
 Le complexe hypothalamo-hypophysaire
est situé à la base du cerveau. Les parois du troisième
ventricule forment l'hypothalamus (1). De sa partie inférieure,
se détache la tige pituitaire (2) qui rejoint l'hypophyse
(3).
Le complexe hypothalamo-hypophysaire
est situé à la base du cerveau. Les parois du troisième
ventricule forment l'hypothalamus (1). De sa partie inférieure,
se détache la tige pituitaire (2) qui rejoint l'hypophyse
(3).
Le complexe hypothalamo-hypophysaire
est situé à la base du cerveau. Les parois du troisième
ventricule forment l'hypothalamus (1). De sa partie inférieure,
se détache la tige pituitaire (2) qui rejoint l'hypophyse
(3).
L'hypophyse est une glande ovoïde, dont la taille est comprise
entre 1,2 et 1,4 cm chez l'homme. Elle est située dans
la selle turcique, petite fossette de la face supérieure
du sphénoïde, incomplètement fermée
par un diaphragme méningé (4).
Voici, en coupe sagittale, les différentes parties du complexe hypothalamo-hypophysaire. A la partie inférieure du troisième ventricule cérébral, les parois de l'hypothalamus fusionnent pour former l'éminence médiane (1). Celle-ci se prolonge par le tuber cinereum ou tige pituitaire (2); son extrémité inférieure dilatée est l'hypophyse postérieure ou posthypophyse (3).
L'antéhypophyse a la forme d'un fer à cheval, elle entoure la posthypophyse. Son bord supérieur est prolongé jusqu'à l'éminence médiane par la pars tuberalis (4). Entre les lobes postérieur (3) et antérieur (5), se trouve le lobe médian (6).
La proximité du chiasma optique (C.O.), situé
en avant et au-dessus de l'hypophyse, explique la fréquence
des troubles oculaires qui accompagnent les tumeurs hypophysaires.
Le lobe antérieur, sa pars tuberalis et le lobe médian forment l'adénohypophyse.
L'éminence médiane, la tige pituitaire et l'hypophyse
postérieure forment ensemble la neurohypophyse.
 L'origine embryonnaire
du complexe hypothalamo-hypophysaire est double. L'hypothalamus
et la neurohypophyse dérivent du neurectoblaste,
coloré en bleu. L'adénohypophyse provient de l'entoblaste,
coloré en rouge (1). Le diencéphale émet
une évagination qui migre caudalement dans le mésenchyme
sous-jacent. En même temps, l'entoblaste stomodéal
forme une vésicule, la poche de Rathke, qui migre
cranialement et s'accole à l'ébauche nerveuse (2).
Après s'être détachée de son épithélium
d'origine (3), l'ébauche entoblastique forme l'adénohypophyse.
Elle entoure partiellement l'ébauche neurectoblastique,
devenue la posthypophyse (4). La paroi antérieure de la
poche de Rathke forme le lobe antérieur. Sa paroi postérieure
forme le lobe médian et sa cavité persiste sous
forme d'une mince fente ou de kystes selon les espèces.
Ce stade terminal est représenté en vue (5) et en
coupe (6) dans ce schéma.
L'origine embryonnaire
du complexe hypothalamo-hypophysaire est double. L'hypothalamus
et la neurohypophyse dérivent du neurectoblaste,
coloré en bleu. L'adénohypophyse provient de l'entoblaste,
coloré en rouge (1). Le diencéphale émet
une évagination qui migre caudalement dans le mésenchyme
sous-jacent. En même temps, l'entoblaste stomodéal
forme une vésicule, la poche de Rathke, qui migre
cranialement et s'accole à l'ébauche nerveuse (2).
Après s'être détachée de son épithélium
d'origine (3), l'ébauche entoblastique forme l'adénohypophyse.
Elle entoure partiellement l'ébauche neurectoblastique,
devenue la posthypophyse (4). La paroi antérieure de la
poche de Rathke forme le lobe antérieur. Sa paroi postérieure
forme le lobe médian et sa cavité persiste sous
forme d'une mince fente ou de kystes selon les espèces.
Ce stade terminal est représenté en vue (5) et en
coupe (6) dans ce schéma.
Voici, en coupe transversale oblique, une hypophyse de chien
à très faible grossissement. La partie centrale,
bleue est la neurohypophyse; elle est prolongée
par la tige pituitaire. L'antéhypophyse est un fer
à cheval qui entoure presque complètement la neurohypophyse.
Une mince fente persiste entre les deux parties principales. Elle
est limitée du côté neurohypophysaire par
une couche épithéliale où les cellules semblent
rangées régulièrement; c'est le lobe intermédiaire.
Dans une coupe transversale d'hypophyse humaine
prélevée à l'autopsie, les dilatations
kystiques du lobe médian sont nombreuses et volumineuses,
elles contiennent une substance amorphe, plissée lors de
la section de la coupe. Le tissu conjonctif y est important.
Le lobe antérieur (A) est formé de cellules groupées
en amas, tandis que le lobe postérieur (P) semble essentiellement
composé de matériel fibrillaire.
 La vascularisation
du complexe hypothalamo-hypophysaire est hautement spécialisée
et adaptée à la régulation de l'hypophyse
par l'hypothalamus. Il existe trois réseaux vasculaires
différents: un réseau hypothalamique pur
(A), un réseau hypothalamo-tubéro-antéhypophysaire
(B) et un réseau post-hypophysaire (C) en relation
avec le précédent.
La vascularisation
du complexe hypothalamo-hypophysaire est hautement spécialisée
et adaptée à la régulation de l'hypophyse
par l'hypothalamus. Il existe trois réseaux vasculaires
différents: un réseau hypothalamique pur
(A), un réseau hypothalamo-tubéro-antéhypophysaire
(B) et un réseau post-hypophysaire (C) en relation
avec le précédent.
Le réseau hypothalamique (A) vascularise l'hypothalamus. Il provient de petites artères issues directement des branches du polygone de Willis. Ses capillaires sont drainés par des veines qui se jettent dans le sinus caverneux.
Le réseau hypothalamo-tubéro-antéhypophysaire (B) vascularise l'éminence médiane, la tige pituitaire et l'antéhypophyse. Il provient des artères hypophysaires supérieures (1), collatérales des carotides ou des artères communicantes postérieures. Les artères hypophysaires supérieures se divisent en deux branches; l'antérieure (2 ), et la postérieure (3) qui se ramifient en un réseau capillaire très dense dans l'éminence médiane et la partie supérieure de la tige pituitaire: le plexus primaire du système porte long (4). La branche antérieure émet aussi l'artère trabéculaire (5) qui descend à la face antérieure de la tige pituitaire, s'enfonce dans l'adénohypophyse et s'anastomose avec une branche de l'artère hypophysaire inférieure (6) pour former le plexus primaire du système porte court (7) dans la partie inférieure de la tige pituitaire. Le plexus primaire du système porte long est drainé par les veines portales longues (8) qui atteignent l'antéhypophyse; les capillaires qui en proviennent forment le plexus antéhypophysaire secondaire (9). Le plexus primaire du système porte court (7) est drainé par les veines portales courtes (10) qui rejoignent le plexus secondaire antéhypophysaire. Celui-ci est drainé à son tour par des veines efférentes (11) qui se jettent dans les sinus de la dure-mère.
Le réseau post-hypophysaire (C) vascularise la neurohypophyse.
Il provient de l'artère hypophysaire inférieure
(6) qui se divise en plusieurs branches pour former un réseau
capillaire dans la neurohypophyse. L'une de ses branches - nous
l'avons mentionné - s'anastomose avec l'artère trabéculaire.
Les capillaires de la neurohypophyse sont drainés par des
veines efférentes (12) qui se jettent également
dans les sinus de la dure-mère.
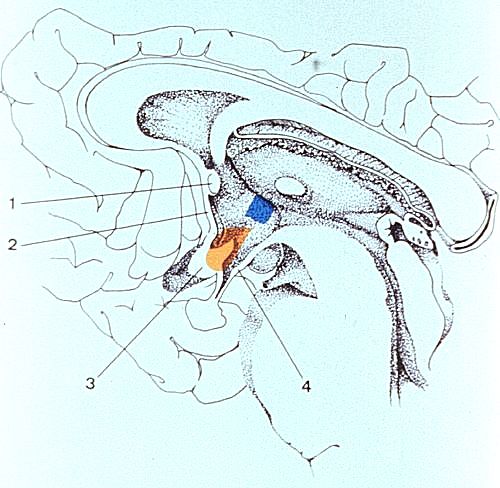 L'hypothalamus
est limité en avant et de haut en bas par la commissure
antérieure (1), la lame terminale (2) et le
chiasma optique (3). Sa limite postérieure est la
fosse interpédonculaire (4). La description complète
de l'hypothalamus n'est pas du ressort de ce cours. Nous nous
limiterons à décrire l'un ou l'autre groupe cellulaire
ou noyau et leurs connexions nerveuses, directement impliqués
dans le fonctionnement du système endocrinien.
L'hypothalamus
est limité en avant et de haut en bas par la commissure
antérieure (1), la lame terminale (2) et le
chiasma optique (3). Sa limite postérieure est la
fosse interpédonculaire (4). La description complète
de l'hypothalamus n'est pas du ressort de ce cours. Nous nous
limiterons à décrire l'un ou l'autre groupe cellulaire
ou noyau et leurs connexions nerveuses, directement impliqués
dans le fonctionnement du système endocrinien.
Nous en avons schématiquement représenté deux: le noyau supraoptique, coloré en orange, et le noyau paraventriculaire coloré en bleu. Chacun a des connexions et un rôle propre.
Les neurones hypothalamiques possédant une fonction endocrine peuvent être divisés en deux systèmes.
Le système magnocellulaire est composé de grandes cellules en relation avec la neurohypophyse.
Le système parvocellulaire est composé de cellules
plus petites, en relation avec l'adénohypophyse.
Les cellules du système magnocellulaire
sont localisées dans des noyaux bien définis; ce
sont les noyaux supraoptiques, situés latéralement
au-dessus du chiasma optique et les noyaux paraventriculaires,
situés plus dorsalement, de chaque côté du
troisième ventricule. Les cellules sont grandes, ont un
noyau volumineux et un cytoplasme intensément coloré
dans cette préparation traitée par la méthode
de Gomori. Il l'est aussi par l'aldéhyde fuchsine. Ces
cellules sont neurosécrétrices: leurs produits
de sécrétion sont contenus dans des grains et associés
à des protéines vectrices, les neurophysines. Ces
grains sont transportés par flux axonal le long de la tige
pituitaire depuis les noyaux hypothalamiques où ils sont
formés, jusqu'à la neurohypophyse où ils
sont stockés, puis sécrétés.
Ces produits de sécrétion sont l'ocytocine
(OCT), liée à la neurophysine I ou A et la vasopressine
ou hormone antidiurétique (ADH) liée à la
neurophysine II ou B. Par immunofluorescence, on a montré
que les noyaux supraoptiques et paraventriculaires participent
conjointement à l'élaboration des deux hormones.
Les neurones du système parvocellulaire
sont regroupés en noyaux bien définis ou disséminés
dans la partie latérale de l'hypothalamus. Ils sont petits,
multipolaires ou fusiformes et possèdent de nombreuses
connexions entre eux. Leur cytoplasme n'est colorable ni par la
méthode de Gomori, ni par l'aldéhyde fuchsine. Leurs
axones sont courts: la plupart se terminent à proximité
du très riche réseau vasculaire de l'éminence
médiane; les autres aboutissent à la paroi du troisième
ventricule.
Ces cellules synthétisent de nombreux petits neuropeptides dont la fonction principale est la régulation de l'adénohypophyse. La localisation de leur production a été étudiée par immunofluorescence. Voici à titre d'exemple, celle de la FSH-RH (follicle stimulating hormone-releasing hormone), de la LH-RH, (luteotropic hormone-releasing hormone) et de la TRH (thyrotropin releasing hormone). Les deux premiers neuropeptides stimulent la sécrétion hypophysaire d'hormones gonadotropes, le troisième stimule la libération d'hormone thyréotrope.
Les neuropeptides sont sécrétés dans les capillaires ou dans le troisième ventricule. Leur présence dans le liquide céphalo-rachidien, leur captation par les cellules épendymaires et leur rapide passage dans les vaisseaux sanguins ont été démontrés. Il est donc possible que le troisième ventricule soit en quelque sorte un lieu de stockage extracellulaire.
Notons que ces neuropeptides ont été retrouvés
en très petites quantités dans d'autres organes
tels que les hémisphères cérébraux,
le tube digestif et même la peau; leur rôle en dehors
de la régulation hypophysaire n'est pas connu.
Les neuropeptides, actuellement isolés ou très probablement présents dans l'hypothalamus, sont groupés en quatre classes. Les deux premières classes règlent l'adénohypophyse, soit en stimulant sa sécrétion (classe 1), soit en l'inhibant (classe 2). Le rôle des deux dernières classes est très mal connu. Certains peptides qui les composent semblent localisés uniquement au niveau du système nerveux central (classe 3), d'autres ont également été retrouvés ailleurs (classe 4). La somatostatine, par exemple, existe aussi dans le pancréas et la thyroïde.
La sécrétion endocrine de l'hypothalamus est principalement réglée par inhibition en retour (feedback) à partir de l'hypophyse ou de glandes périphériques. Elle l'est en outre par le système monoaminergique et le système limbique.
Le système monoaminergique agit sur les systèmes
magno et parvocellulaires par des médiateurs libérés
dans plusieurs régions hypothalamiques mais surtout dans
l'éminence médiane: la dopamine, la noradrénaline
et la sérotonine. Ces médiateurs agiraient
aussi directement sur l'hypophyse après avoir été
libérés dans le système porte hypophysaire.
La sécrétion de prolactine serait ainsi inhibée
par la dopamine.
Le système limbique intervient surtout sur le système
parvocellulaire; cette régulation est très mal connue.
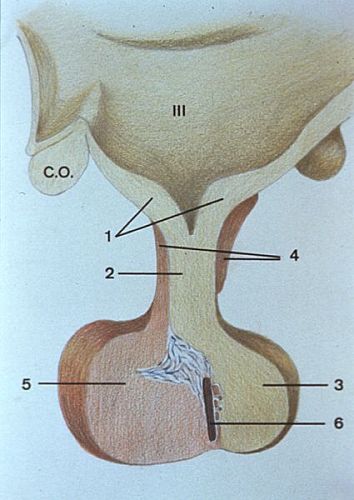 La neurohypophyse
comprend l'éminence médiane (1), considérée
par certains comme une part de l'hypothalamus, la tige pituitaire
(2) et la posthypophyse (3).
La neurohypophyse
comprend l'éminence médiane (1), considérée
par certains comme une part de l'hypothalamus, la tige pituitaire
(2) et la posthypophyse (3).
L'adénohypophyse est formée du lobe tubéral
ou pars tuberalis (4), du lobe antérieur
ou pars distalis (5), le plus important en volume,
et du lobe médian (6). Elle est composée
de cellules épithéliales endocrines à sécrétion
protéique. Il en existe plusieurs types, répartis
en proportions inégales dans les différentes régions.
Les cellules épithéliales
des lobes antérieur et tubéral sont disposées
en îlots ou en cordons séparés
par des capillaires; leur noyau est arrondi, il contient
1 à 2 nucléoles. Leur cytoplasme est granulaire.
Dans cette préparation, le cytoplasme de certaines cellules
est acidophile et donc coloré en rouge; le cytoplasme
d'autres cellules est basophile et coloré en bleu.
De rares cellules de petite taille, dites chromophobes
ont un cytoplasme incolore qui apparaît comme un halo clair
autour du noyau. L'une d'elles est indiquée par une flèche.
Chez l'homme, la disposition en îlots
est plus marquée parce que le tissu conjonctif est plus
développé.
Il est possible d'établir une corrélation entre
l'aspect histologique habituel des cellules et la nature de leur
sécrétion. Les cellules acidophiles sécrètent
soit l'hormone de croissance ou GH, soit la prolactine.
Les cellules basophiles sécrètent soit l'hormone
thyréotrope ou TSH, soit les gonadotrophines LH
et FSH, soit l'hormone corticotrope ou ACTH. Le rôle
des cellules chromophobes n'est pas clair: il semble qu'elles
soient, chez l'homme en tout cas, des cellules indifférenciées
ou dégranulées.
Des colorations plus spécifiques permettent de différencier
les cellules à GH et à prolactine. Ainsi, dans une
coupe colorée selon la méthode tétrachromique de Herlant,
les cellules à GH sont oranges et les cellules à
prolactine sont rouges; les cellules basophiles, elles,
restent bleues. Il faut cependant admettre que la distinction
entre les teintes cytoplasmiques n'est pas toujours chose aisée.
Avec l'aldéhyde fuchsine,
le cytoplasme des cellules à TSH (T) est coloré
en mauve tandis que celui des cellules à gonadotrophines
(G) est coloré en rouge.
Pour localiser une hormone de façon précise dans
un type cellulaire déterminé, il faut utiliser des
techniques immunologiques. Voici à titre d'exemple,
la détection des cellules sécrétant de la GH
révélée par double immunofluorescence
dans l'hypophyse d'une souris très jeune.
Et voici la localisation des cellules à ACTH
chez le même animal.
L'ultrastructure cellulaire et surtout l'aspect des
grains permet aussi d'identifier les cellules et leur sécrétion.
La cellule qui sécrète l'hormone de croissance
est ronde; son réticulum endoplasmique rugueux est bien
développé, les grains de sécrétion
sont ronds et nombreux, leur taille est comprise entre 350 et
400 nanomètres.
Les grains de la cellule qui sécrète la prolactine
sont différents. Leur forme est irrégulière,
leur taille varie de 550 à 600 nanomètres.
La cellule qui sécrète l'ACTH
est la plus typique. Elle est allongée; ses grains sont
peu nombreux, ont un diamètre variant entre 200 et 250
nanomètres, et sont disposés régulièrement
le long de la membrane plasmique.
Voici un résumé des caractères morphologiques
et de la fonction des cellules antéhypophysaires.
Les cellules acidophiles sécrétant la GH sont colorables à l'orange G qui masque leur affinité pour l'érythrosine. Elles sont rondes, leur réticulum endoplasmique rugueux est très développé. Les grains de sécrétion sont très nombreux, modérément denses, ronds et leur diamètre est compris entre 350 et 400 nanomètres. Elles représentent 50% du nombre total des cellules antéhypophysaires chez l'adulte. Leur proportion est plus grande chez l'enfant. Un facteur hypothalamique, le GH-RH, l'hypoglycémie et le stress augmentent leur sécrétion. La somatostatine l'inhibe. L'effet de l'hormone de croissance se résume en trois mots: elle est anabolisante, hyperglycémiante et lipotrope.
Les cellules acidophiles sécrétant la prolactine sont colorables en rouge par l'érythrosine mais pas à l'orange G. Leur cytoplasme est riche en réticulum endoplasmique rugueux; leurs grains sont irréguliers, très denses et ont un diamètre compris entre 550 et 600 nanomètres. Elles représentent 10 à 20% des cellules antéhypophysaires. Au cours de la grossesse, elles augmentent en nombre et en taille. La sérotonine, les catécholamines et l'acide gamma-amino-butyrique stimulent leur sécrétion. Le PIF et la dopamine l'inhibent.
La prolactine stimule le développement de la glande mammaire
pendant la grossesse, entretient la sécrétion lactée
pendant l'allaitement et intervient dans le cycle menstruel.
Quatre hormones, la TSH, la LH, la FSH et l'ACTH sont sécrétées par les cellules basophiles. La TSH, la LH et la FSH ont toutes les trois la même chaîne alpha et chacune une chaîne bêta spécifique.
Les cellules basophiles qui sécrètent la TSH sont PAS positives et colorables à l'aldéhyde fuchsine. Elles sont polygonales, leur réticulum endoplasmique rugueux est réduit, leurs grains, ronds et peu denses, sont les plus petits de l'adénohypophyse (150 nanomètres). Elles représentent 5% des cellules antéhypophysaires; l'hypothyroïdie augmente leur taille et leur nombre. La TRH ou thyrolibérine stimule leur sécrétion tandis que les hormones thyroïdiennes l'inhibent. La TSH a un effet tonique et trophique sur la thyroïde.
Les cellules basophiles qui sécrètent les gonadotrophines sont PAS négatives et ne sont pas colorées en mauve par l'aldéhyde fuchsine. Elles sont volumineuses, riches en réticulum endoplasmique rugueux aux citernes dilatées. Leurs grains sont peu nombreux et leur diamètre varie entre 100 à 300 nanomètres. Selon certains, la LH et la FSH seraient sécrétées par des cellules différentes, mais l'immunohistochimie a démontré la présence des deux hormones dans une même cellule. La sécrétion de LH est contrôlée par la LH-RH; celle de FSH par la FSH-RH. Les stéroïdes sexuels exercent sur cette sécrétion une régulation très complexe.
Chez le mâle, la FSH stimule la spermatogenèse; la LH stimule la synthèse de la testostérone par les cellules interstitielles du testicule.
Chez la femelle, la FSH stimule la croissance du follicule ovarien tandis que la LH contrôle l'ovulation et la lutéinisation.
Les cellules basophiles qui sécrètent l'ACTH
sont PAS positives mais ne sont pas colorables à l'aldéhyde
fuchsine. Elles sont allongées; leurs grains ronds dont
le diamètre varie entre 200 et 250 nanomètres sont
disposés de manière caractéristique le long
de la membrane plasmique. Elles forment 20% des cellules adénohypophysaires,
mais sont surtout concentrées dans le lobe tubéral.
Le CRH hypothalamique stimule la sécrétion d'ACTH;
les glucocorticoïdes surrénaliens l'inhibent. L'ACTH
contrôle les zones fasciculaire et réticulaire de
la surrénale. Elle présente de nombreuses similitudes
avec la MSH.
La taille du lobe médian
varie selon les espèces et l'age. Chez l'enfant, il est
volumineux; chez l'adulte, il ne représente que 2% de l'hypophyse
et est formée de kystes et de quelques cellules
basophiles, sécrétant la MSH.
Les similitudes morphologiques entre les cellules à ACTH
et à MSH s'expliquent par la façon dont ces hormones
sont synthétisées. Toutes deux dérivent d'un
précurseur commun. En suivant dans ce schéma,
depuis son extrémité N terminale jusqu'à
son extrémité C terminale, on retrouve successivement
le signal de sécrétion, puis un peptide de 16.000
daltons dont le rôle est inconnu, puis l'ACTH (39
acides aminés) et enfin la bêta lipotrophine (91
acides aminés). L'ACTH peut être scindé en
alpha-MSH (13 acides aminés) et en CLIP (11 acides aminés).
La bêta lipotrophine peut être scindée en gamma
lipotrophine (58 acides aminés) et en bêta endorphine
(30 acides aminés). Les acides aminés 41 à
55 de la gamma lipotrophine forment la bêta-MSH et les acides
aminés 63 à 67 la métenképhaline.
Alpha et bêta-MSH ont un effet stimulant sur les mélanocytes
cutanés et provoquent l'apparition d'une pigmentation brunâtre.
L'ACTH a le même effet puisque ses 13 premiers acides aminés
correspondent à l'alpha-MSH.
Réexaminons la coupe d'hypophyse de chien
à faible grossissement. La partie bleue est la neurohypophyse.
Dans celle-ci et de gauche à droite, se retrouvent une
partie globuleuse, l'hypophyse postérieure, une région
étroite, située entre les deux extrémités
de l'hypophyse antérieure, la tige pituitaire et l'éminence
médiane. Celle-ci est bifide dans sa partie supérieure;
sa fente est le troisième ventricule, limité par
un épithélium cubique, l'épithélium
épendymaire.
Examinons cette coupe au grossissement moyen. La neurohypophyse
est composée d'un matériel fibrillaire de densité
variable. Quelques vaisseaux, reconnaissables aux globules rouges
colorés en orange, sont disséminés dans le
tissu. Seuls les noyaux brunâtres des cellules tranchent
nettement sur le fond bleu. Plusieurs taches rondes de forme et
de taille variables sont dispersées dans le matériel
fibrillaire: ce sont les corps de Hering.
A fort grossissement, les corps de Hering
sont aisément reconnaissables dans une coloration trichromique
à droite, ou tétrachromique à gauche. Ils
sont ronds ou ovalaires, coloriés en brun ou en bleu. Leur
contenu est finement granulaire.
Les corps de Hering résultent de l'accumulation des produits de sécrétion dans les terminaisons dilatées des axones hypothalamiques. La vasopressine et l'ocytocine y sont contenues dans des grains enveloppés par une membrane, leur diamètre mesure environ 150 nanomètres.
La terminaison axonique est enveloppée par les expansions
cytoplasmiques de cellules gliales spécialisées.
Le produit de sécrétion de la neurohypophyse, élaboré
par les neurones hypothalamiques du système magnocellulaire,
est transporté dans le corps de Hering par flux axonal,
à la vitesse de 1 à 2 cm à l'heure. Dans
cette micrographie,
les axones ont été coupés longitudinalement.
Les grains de sécrétion y sont disposés le
long de microtubules qui sont impliqués dans leur transport.
Les cellules endothéliales
sont reconnaissables à leur noyau dense, allongé
en coupe longitudinale, petit et rond en coupe transversale. Leur
cytoplasme limite la lumière des vaisseaux visible lorsqu'elle
contient des globules rouges colorés en orange. Lorsque
la lumière vasculaire est collabée, la cellule endothéliale
est une fine ligne bleue plus foncée. Ces capillaires sont
fenestrés.
La cellule gliale
est la cellule la plus fréquente de la neurohypohyse. En
microscopie optique, on la reconnaît à son noyau
volumineux dont l'euchromatine est importante. En microscopie
électronique, on peut observer ses expansions entourant
incomplètement le corps de Hering. Une mince fente persiste
entre les deux membranes plasmiques. On ignore si la cellule gliale
joue un rôle actif dans la régulation de la sécrétion
des hormones à partir des corps de Hering.
Les cellules de la microglie
sont petites, leur noyau est dense, arrondi ou ovalaire. Dans
les coupes classiques, on les repère difficilement; leur
petit noyau est enveloppé d'une fine bande cytoplasmique,
plus foncée que les corps de Hering. En microscopie électronique,
elles sont reconnaissables parce qu'elles possèdent de
nombreux et volumineux lysosomes. Les cellules de la microglie
sont l'équivalent dans le système nerveux central
des macrophages trouvés dans les autres tissus.
Les deux produits de la neurohypohyse sont l'ocytocine et la vasopressine, peptides de neuf acides aminés dont deux seulement diffèrent; ceci explique leur activité biologique croisée.
L'ocytocine provoque les contractions du muscle utérin au cours de l'accouchement et l'injection du lait au cours de la lactation. Sa sécrétion est stimulée par un réflexe neurohormonal dont l'origine se trouve dans les terminaisons nerveuses sensitives du vagin, du col utérin et de l'aréole mammaire.
La vasopressine agit sur le muscle lisse des vaisseaux
et réduit la diurèse en augmentant la perméabilité
des canaux collecteurs des reins. La sécrétion de
l'ADH est stimulée par l'hypertonicité du plasma
qui active des cellules hypothalamiques spécialisées,
dites osmoréceptrices. L'absence d'ADH provoque une diurèse
importante, appelée diabète insipide, que celui
qui en souffre compense par une boisson abondante.
La prochaine leçon sera consacrée
à la thyroïde, à la surrénale et aux
cellules endocrines des gonades.