Le système digestif (Cours 7)
Les glandes salivaires sont des glandes exocrines
annexées à la cavité buccale. Il existe des
glandes salivaires microscopiques ou accessoires et des glandes
salivaires macroscopiques ou principales. L'ensemble des sécrétions
de ces glandes constitue la salive. Nous terminerons en précisant
sa composition, ses fonctions et le contrôle de sa sécrétion.
Les glandes salivaires accessoires
sont éparpillées dans le chorion des muqueuses jugales,
labiales, linguale, sublinguale, palatine et pharyngée.
Elles sont très petites, de 1 à 3 mm de diamètre
et constituées de quelques acini drainés par un
canal excréteur court, non ou peu ramifié. Généralement
les acini, muqueux et séreux, sont mélangés.
Il existe cependant des glandes séreuses pures (A), situées
notamment dans le fond du sillon entourant les papilles circonvallariées
de la langue où elles sont appelées glandes de
von Ebner, et des glandes muqueuses pures (B) disséminées
au niveau du palais et de la base de la langue.
La sécrétion des glandes salivaires accessoires
est continue. Au niveau de la langue, elle rince les sillons des
papilles, imbibe les bourgeons gustatifs et contribue ainsi aux
qualités discriminatives de la gustation.
 Les trois paires de glandes salivaires principales
sont disposées symétriquement dans les parois de
la cavité buccale où leur produit de sécrétion
est déversé par des canaux excréteurs distincts.
Elles sont anatomiquement individualisées et sont appelées
: glande parotide (1), glande sousmaxillaire
(2) et glande sublinguale (3). Leur sécrétion
est discontinue, induite par des stimuli mécaniques, thermiques,
chimiques, psychiques ou olfactifs. Au début de leur développement
embryologique, ces glandes sont des expansions pleines de l'épithélium
buccal. Les structures sécrétrices se forment après
le système canalaire, à partir des extrémités
des canaux les plus fins. La morphogenèse ne sera définitive
qu'après la naissance, pour les éléments
séreux, et un peu avant, pour les éléments
muqueux. Le mésenchyme adjacent et le système nerveux,
surtout orthosympathique, jouent un rôle important dans
cette morphogenèse.
Les trois paires de glandes salivaires principales
sont disposées symétriquement dans les parois de
la cavité buccale où leur produit de sécrétion
est déversé par des canaux excréteurs distincts.
Elles sont anatomiquement individualisées et sont appelées
: glande parotide (1), glande sousmaxillaire
(2) et glande sublinguale (3). Leur sécrétion
est discontinue, induite par des stimuli mécaniques, thermiques,
chimiques, psychiques ou olfactifs. Au début de leur développement
embryologique, ces glandes sont des expansions pleines de l'épithélium
buccal. Les structures sécrétrices se forment après
le système canalaire, à partir des extrémités
des canaux les plus fins. La morphogenèse ne sera définitive
qu'après la naissance, pour les éléments
séreux, et un peu avant, pour les éléments
muqueux. Le mésenchyme adjacent et le système nerveux,
surtout orthosympathique, jouent un rôle important dans
cette morphogenèse.
Les glandes salivaires principales
comprennent des éléments sécréteurs
regroupés en grappes (A) et constitués d'acini séreux,
muqueux ou séromuqueux et un système canalaire
ramifié (B), partagé en différents segments.
Les canaux intralobulaires comprennent les canaux intercalaires
et les canaux striés. Chaque canal intercalaire draine
un acinus, chaque canal strié draine un lobule. Plusieurs
canaux striés confluent vers un canal interlobulaire qui
aboutit au canal excréteur principal. La capsule conjonctive
de la glande émet des cloisons qui la partagent en lobes
et en lobules et renferment des vaisseaux sanguins, des nerfs
et les canaux excréteurs interlobulaires et principal.
L'aspect histologique des cellules muqueuses, des cellules séreuses,
des cellules myoépithéliales, de la membrane basale
et des acini a été décrit dans le cours d'histologie
générale. Les acini mixtes,
séromuqueux possèdent à la fois des cellules
séreuses et des cellules muqueuses agencées de telle
sorte que la paroi de l'acinus est tapissée par les cellules
muqueuses et que son extrémité borgne est coiffée
par les cellules séreuses disposées en croissants,
dits de Gianuzzi. La sécrétion de ces cellules
séreuses est drainée par de fins canalicules qui
serpentent entre les cellules muqueuses et se jettent dans la
lumière de la glande. Ces canalicules ne sont pas visibles
à ce grossissement.
Grâce à des colorants spécifiques du mucus,
tel que le mucicarmin, on peut distinguer les cellules muqueuses
colorées en rouge et disposées le long de la lumière
glandulaire et les cellules séreuses disposées
en croissant autour des cellules muqueuses.
La lumière des canaux intercalaires
(A) est étroite et bordée par des cellules aplaties;
celle des canaux striés (B) qui leur font suite
est plus large et bordée de cellules épithéliales
pyramidales, unies à leur pôle apical par des
complexes de jonction. Leur pôle basal est strié.
Cette striation basale
est due à de nombreuses mitochondries verticales, insérées
entre des invaginations parallèles de la membrane plasmique.
Le pôle cellulaire apical est, pour sa part, hérissé
de nombreuses microvillosités. Les cellules des
canaux striés ressemblent aux cellules des tubes contournés
proximaux du rein et sont comme elles impliquées dans le
transport actif des électrolytes, notamment du sodium
qui y est résorbé. La salive produite par les glandes
pourvues de canaux striés devient ainsi hypotonique
tandis que celle produite par les glandes dépourvues de
canaux striés est isotonique.
Le calibre des canaux interlobulaires
(A) et principaux (B) croît au fur et à mesure que
l'on se rapproche de la cavité buccale et leur épithélium
cylindrique simple est progressivement modifié pour
devenir pavimenteux stratifié près de l'orifice
buccal. Cet épithélium s'enrichit par endroits de
cellules caliciformes et de cellules myoépithéliales.
La salive ne semble pas y subir de modifications quantitatives
notables.
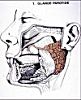 La parotide est
la plus volumineuse des glandes salivaires. Elle occupe une loge
située devant l'oreille, entre la mastoïde, la branche
montante de la mandibule et l'arcade zygomatique sous laquelle
elle se prolonge. Le canal excréteur, ou canal de Stenon,
part de cette portion souszygomatique, chemine dans la joue
et s'ouvre dans la cavité buccale à hauteur de la
deuxième prémolaire supérieure.
La parotide est
la plus volumineuse des glandes salivaires. Elle occupe une loge
située devant l'oreille, entre la mastoïde, la branche
montante de la mandibule et l'arcade zygomatique sous laquelle
elle se prolonge. Le canal excréteur, ou canal de Stenon,
part de cette portion souszygomatique, chemine dans la joue
et s'ouvre dans la cavité buccale à hauteur de la
deuxième prémolaire supérieure.
La parotide est enveloppée par une capsule fibreuse
de laquelle se détachent de nombreux septa qui divisent
la glande en lobes et en lobules. Le tissu conjonctif de cette
armature fibreuse renferme des vaisseaux, des nerfs et des adipocytes
dont le nombre croît avec l'âge. Les canaux interlobulaires
y cheminent également.
Le parenchyme de la parotide
est constitué d'acini séreux. Leur lumière
punctiforme est délimitée par une seule couche de
cellules pyramidales. Les cellules séreuses renferment
dans leur pôle apical des grains de zymogène,
dissous, dans ce casci, au cours de la préparation
du tissu. Ils sont constitués de protéines libérées
sous forme d'enzymes dans la sécrétion salivaire.
Le pôle basal de ces cellules contient le noyau et est riche
en ribosomes.
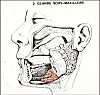 La glande sousmaxillaire
est située dans le plancher de la bouche sous le corps
de la mandibule, entre celuici et les muscles. Le canal
excréteur, ou canal de Wharton, s'ouvre sur le côté
du frein de la langue.
La glande sousmaxillaire
est située dans le plancher de la bouche sous le corps
de la mandibule, entre celuici et les muscles. Le canal
excréteur, ou canal de Wharton, s'ouvre sur le côté
du frein de la langue.
La sousmaxillaire
est entourée par une capsule fibreuse bien délimitée.
La plupart des acini sont purement séreux, quelquesuns
sont muqueux mais le plus souvent les cellules muqueuses
forment avec les cellules séreuses des acini mixtes
(flèches). Les canaux de la glande sont aussi bien développés
que ceux de la parotide; les canaux intercalaires sont cependant
plus courts et par conséquent se voient moins souvent dans
les coupes histologiques, tandis que les canaux striés
sont plus longs et donc mieux visibles. Les lobules de la glande
sont séparés les uns des autres par du tissu conjonctif
riche en adipocytes.
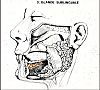 La glande sublinguale
est allongée en dedans de la glande sousmaxillaire,
directement sous la muqueuse du plancher buccal. Cette glande
est en fait un agglomérat de huit à vingt petites
glandes, s'ouvrant individuellement dans le plancher buccal. Le
canal le plus important ou canal de Bartholin possède
un orifice proche mais indépendant du canal de Wharton.
La glande sublinguale
est allongée en dedans de la glande sousmaxillaire,
directement sous la muqueuse du plancher buccal. Cette glande
est en fait un agglomérat de huit à vingt petites
glandes, s'ouvrant individuellement dans le plancher buccal. Le
canal le plus important ou canal de Bartholin possède
un orifice proche mais indépendant du canal de Wharton.
Il n'y a pas de réelle capsule autour de la sublinguale
mais les septa conjonctifs sont plus importants que dans la parotide
et la sousmaxillaire. Dans la glande sublinguale,
les acini purement séreux sont rares, les quelques
cellules séreuses retrouvées forment des croissants
de Gianuzzi; la plupart des acini sont donc purement muqueux.
Les canaux intralobulaires, tant intercalaires que striés,
sont très courts. Les canaux principaux sont nombreux,
droits et courts.
L'innervation sensitive des glandes salivaires principales
est assurée par le nerf trijumeau. L'innervation
effectrice est assurée par le système
autonomique via le nerf glossopharyngien et le ganglion otique
pour la glande parotide et via la corde du tympan et le ganglion
mandibulaire pour la sousmaxillaire et la sublinguale.
La vascularisation sanguine est assurée par l'artère
faciale et la carotide externe pour la glande parotide, les artères
faciale et linguale pour la glande sous-maxillaire, les artères
sublinguale et sousmentonnière pour la glande sublinguale.
Les capillaires sont disposés en deux réseaux réunis
par des veines portes. Le premier réseau est disposé
autour des canaux et le second autour des acini. Le flux sanguin
circule donc en sens inverse du flux salivaire, ce qui
permet une concentration de la salive.
Ce tableau résume les caractéristiques les plus
importantes des glandes salivaires principales.
- La glande parotide est à prédominance
séreuse. Les canaux intercalaires et striés sont
bien visibles. Ils aboutissent au canal de Stenon. Le tissu conjonctif
est bien développé et forme une capsule nette, riche
en cellules adipeuses.
- La glande sousmaxillaire est composée
d'acini séreux et séromuqueux. Les canaux intercalaires
sont courts et donc moins visibles. Les canaux striés sont
longs. Ils aboutissent au canal de Wharton. Le tissu conjonctif
est bien développé.
- La glande sublinguale est à prédominance
muqueuse. Les canaux intercalaires et striés sont courts.
Ils aboutissent à 8 ou 10 canaux excréteurs dont
celui de Bartholin. Il n'existe pas de capsule conjonctive propre
mais les septa sont bien développés.
La salive est l'ensemble des sécrétions de toutes
les glandes salivaires. Son volume quotidien est d'un litre à
un litre et demi et les glandes sousmaxillaires assurent
les deux tiers de cette sécrétion. La sécrétion
des glandes salivaires principales est discontinue,
consécutive à des stimuli principalement lies à
l'absorption d'aliments. La sécrétion des glandes
salivaires accessoires est continue mais son volume quotidien
est infime par rapport au volume salivaire total. La salive est
une solution aqueuse d'immunoglobulines, d'enzymes
tels que le lysozyme, l'amylase et la maltase, d'autres protéines,
de sels minéraux et de métaux lourds.
Dans la cavité buccale, la salive s'enrichit en cellules
pavimenteuses desquamées, en lymphocytes, en polynucléaires
et en éléments bactériens appartenant à
la flore commensale.
La salive assure l'humidification constante et la lubrification
des muqueuses buccale et labiales, le rinçage de la bouche
et l'élimination des débris alimentaires et cellulaires
afin d'éviter la fermentation de ceuxci et la pullulation
des bactéries toujours présentes dans la bouche.
Elle intervient dans la digestion en ramollissant les aliments
ce qui facilite leur déglutition et permet une meilleure
perception des saveurs par les bourgeons gustatifs. Le rôle
des enzymes salivaires est évanescent car elles sont neutralisées
par le pH acide de la cavité gastrique.
La sécrétion de la salive intervient dans le maintien
de la balance hydrique car la déshydratation tarit
la sécrétion salivaire créant la sensation
de soif. Outre les électrolytes, les métaux lourds
sont excrétés par la salive. Les glandes salivaires
concentrent les iodures et le calcium.
Le contrôle de la sécrétion salivaire est
assure en grande partie par les systèmes nerveux orthosympathique
et parasympathique, notamment sous l'effet de stimuli locaux,
gustatifs, olfactifs, irritatifs des muqueuses buccale, oesophagienne
ou gastrique, ou de stimuli psychiques. La stimulation expérimentale
du système orthosympathique entraîne la sécrétion
d'une salive épaisse et muqueuse, tandis que la
stimulation parasympathique provoque la sécrétion
d'une salive fluide et abondante.
 Le pancréas
se forme à partir de deux ébauches, l'une ventrale
(1) contiguë à l'ébauche hépatique (2)
et l'autre dorsale (3). L'une et l'autre sont des excroissances
endodermiques duodénales (4). L'ébauche
dorsale (3) dont la croissance est plus importante, formera
la majeure partie de la glande et est centrée sur un canal
excréteur, le canal de Santorini (A). L'ébauche
ventrale suit le duodénum au cours de sa rotation puis
s'accole et fusionne avec la partie inférieure de la portion
juxtaduodénale de l'ébauche dorsale. Cette
ébauche ventrale deviendra la moitié inférieure
de la tête du pancréas et est drainée par
le canal de Wirsung (B) qui, chez 90% des individus, fusionne
avec le canal de Santorini et devient le canal excréteur
unique.
Le pancréas
se forme à partir de deux ébauches, l'une ventrale
(1) contiguë à l'ébauche hépatique (2)
et l'autre dorsale (3). L'une et l'autre sont des excroissances
endodermiques duodénales (4). L'ébauche
dorsale (3) dont la croissance est plus importante, formera
la majeure partie de la glande et est centrée sur un canal
excréteur, le canal de Santorini (A). L'ébauche
ventrale suit le duodénum au cours de sa rotation puis
s'accole et fusionne avec la partie inférieure de la portion
juxtaduodénale de l'ébauche dorsale. Cette
ébauche ventrale deviendra la moitié inférieure
de la tête du pancréas et est drainée par
le canal de Wirsung (B) qui, chez 90% des individus, fusionne
avec le canal de Santorini et devient le canal excréteur
unique.
Le pancréas
mesure 20 à 25 cm de long et pèse entre 65 et 160
grammes, 99% de sa masse étant constitués par sa
portion exocrine. Sa configuration est telle qu'il a été
comparé à un crochet ou à un marteau. Il
comporte trois segments : la tête accolée
à la concavité duodénale, le corps
et la queue adjacente au hile splénique. Lorsqu'il
est frais, le pancréas est rose pâle et est recouvert
par un très fin tissu fibreux insuffisant pour constituer
une capsule véritable et pour masquer sa lobulation. La
vascularisation artérielle est assurée par les artères
gastroduodénale, mésentérique supérieure
et splénique. La circulation veineuse aboutit au système
porte hépatique. Les nerfs suivent les artères;
ils proviennent du plexus coeliaque (fibres nerveuses orthosympathiques
amyélinisées) et du pneumogastrique (fibres
nerveuses parasympathiques myélinisées). Le drainage
lymphatique est assure par les différentes chaînes
voisines.
Le tissu conjonctif du pancréas
est pauvre en fibres et peu abondant; il forme des septa qui divisent
l'organe en lobules et une enveloppe ténue qui recouvre
l'organe et est tapissée à la face ventrale par
le péritoine. Les septa sont parcourus par les vaisseaux
sanguins et lymphatiques, par les nerfs et par les canaux excréteurs
(A); ils contiennent quelques ganglions lymphatiques, de petits
ganglions nerveux et des corpuscules de VaterPaccini (B).
Le tissu conjonctif se condense davantage autour des canaux excréteurs
interlobulaires et principaux. La quantité d'adipocytes
augmente avec l'age.
L'acinus séreux pancréatique
se distingue de l'acinus des glandes salivaires par des contours
plus irréguliers, l'absence de cellules myoépithéliales
et la présence de cellules centro-acineuses. L'aspect
et la taille des acini et des cellules séreuses sont fonction
de leur degré d'activité. Le tissu conjonctif contigu
est lâche et renferme des capillaires et des terminaisons
nerveuses libres qui entrent en contact avec le pôle basal
des cellules sécrétrices.
Les cellules séreuses,
de forme pyramidale, ont un pôle basal parce qu'il est riche
en ergastoplasme, un noyau rond, situé dans la moitié
basale de la cellule, et un pôle apical qui renferme des
grains de zymogène. Les cellules centro-acineuses
appartiennent aux voies excrétrices et délimitent
en fait un prolongement du canal intercalaire au centre même
de l'acinus. Les cellules centro-acineuses ont un noyau central
et rond, pauvre en hétérochromatine et un cytoplasme
pâle peu important, pauvre en organites.
Le canal intercalaire
(A) commence donc le plus souvent au sein même de l'acinus,
par les cellules centro-acineuses; parfois, il fait simplement
suite à l'acinus sans s'y prolonger. Le canal intercalaire
est long et étroit, bordé d'un épithélium
simple et aplati. Il se jette dans un canal intralobulaire
désigné par une flèche en (B) dont l'épithélium
est cubique. Ce canal intralobulaire, très court, n'est
pas strié. L'épithélium des canaux interlobulaires
(C) est d'abord cubique puis cylindrique simple; il contient quelques
cellules caliciformes.
Les canaux interlobulaires
s'abouchent à angle plus ou moins droit au canal de
Wirsung qui parcourt la glande sur toute sa longueur, l'ensemble
présentant une architecture en arête de poisson.
Le canal de Wirsung augmente de calibre de la queue du pancréas
vers la tête ou il rejoint le canal cholédoque
avec lequel il s'ouvre dans la lumière duodénale
au niveau d'un orifice unique, l'ampoule de Vater, entourée
par le sphincter musculaire lisse d'Oddi.
Parfois les deux canaux s'abouchent au duodénum séparément.
L'épithélium du canal de Wirsung est cylindrique
pluristratifié et repose sur un chorion riche en fibres
de collagène et en fibres élastiques; on y observe,
prés de l'orifice duodénal, quelques glandes muqueuses
et des cellules musculaires lisses.
L'histophysiologie du pancréas exocrine, par laquelle nous
terminerons cette leçon, s'intéresse à la
sécrétion du suc pancréatique et à
la régulation de cette sécrétion. Un
pancréas adulte sécrète quotidiennement dans
la lumière duodénale environ deux litres et demi
d'un suc aqueux incolore et alcalin, riche en électrolytes,
en bicarbonates et en enzymes. Ce liquide alcalin
contribue à neutraliser, par sa richesse en ions
bicarbonates, l'acidité du chyme gastrique avant son entrée
dans le jéjunoiléon. L'eau et les bicarbonates
proviennent en grande partie des cellules centro-acineuses
et des canaux excréteurs.
Les enzymes du suc pancréatique dégradent
les protéines, les lipides, les hydrates de carbone et
les acides nucléiques du bol alimentaire; elles comptent
des peptidases, une lipase, des estérases, une amylase
et des nucléases. Exceptées la lipase et l'amylase,
toutes ces enzymes sont sécrétées sous
forme inactive et sont activées dans la lumière
intestinale, ce qui protège le parenchyme pancréatique
de l'autodigestion. A titre d'exemple retenons que l'entérokinase,
sécrétée par la muqueuse duodénale,
active le trypsinogène en trypsine qui ellemême
active le chymotrypsinogène en chymotrypsine.
La sécrétion enzymatique
est mérocrine. Elle résulte de l'exocytose des grains
de zymogène par les cellules séreuses dans la lumière
de l'acinus dont le volume est ainsi accru, tandis que les cellules
raccourcissent.
La régulation de cette sécrétion pancréatique
est hormonale et nerveuse. La sécrétion
pancréatique est minime dans des conditions basales, elle
s'accroît considérablement lors des repas. Cette
activation est principalement contrôlée par des hormones
sécrétées par les cellules endocrines
des muqueuses pylorique et duodénale, atteignant par
voie sanguine les canaux et acini pancréatiques. La sécrétine
est libérée par la muqueuse duodénale au
contact du chyme acide et active la sécrétion de
l'eau et des bicarbonates par les cellules canalaires. La pancréozymine,
la cholecystokinine et la gastrine sont sécrétées,
les deux premières par la muqueuse duodénale, la
troisième par la muqueuse pylorique, en présence
de produits de dégradation des protéines. Elles
stimulent la libération des enzymes dans le suc pancréatique
ainsi que l'évacuation concomitante de bile.
La régulation nerveuse de la sécrétion
pancréatique paraît évidente mais est mal
expliquée. Expérimentalement, la stimulation électrique
ou psychique des pneumogastriques augmente la teneur en enzymes
de cette sécrétion. En fait, ce contrôle nerveux
pourrait être subordonné à la régulation
hormonale.
La prochaine leçon sera consacrée
au foie.
 Les trois paires de glandes salivaires principales
sont disposées symétriquement dans les parois de
la cavité buccale où leur produit de sécrétion
est déversé par des canaux excréteurs distincts.
Elles sont anatomiquement individualisées et sont appelées
: glande parotide (1), glande sousmaxillaire
(2) et glande sublinguale (3). Leur sécrétion
est discontinue, induite par des stimuli mécaniques, thermiques,
chimiques, psychiques ou olfactifs. Au début de leur développement
embryologique, ces glandes sont des expansions pleines de l'épithélium
buccal. Les structures sécrétrices se forment après
le système canalaire, à partir des extrémités
des canaux les plus fins. La morphogenèse ne sera définitive
qu'après la naissance, pour les éléments
séreux, et un peu avant, pour les éléments
muqueux. Le mésenchyme adjacent et le système nerveux,
surtout orthosympathique, jouent un rôle important dans
cette morphogenèse.
Les trois paires de glandes salivaires principales
sont disposées symétriquement dans les parois de
la cavité buccale où leur produit de sécrétion
est déversé par des canaux excréteurs distincts.
Elles sont anatomiquement individualisées et sont appelées
: glande parotide (1), glande sousmaxillaire
(2) et glande sublinguale (3). Leur sécrétion
est discontinue, induite par des stimuli mécaniques, thermiques,
chimiques, psychiques ou olfactifs. Au début de leur développement
embryologique, ces glandes sont des expansions pleines de l'épithélium
buccal. Les structures sécrétrices se forment après
le système canalaire, à partir des extrémités
des canaux les plus fins. La morphogenèse ne sera définitive
qu'après la naissance, pour les éléments
séreux, et un peu avant, pour les éléments
muqueux. Le mésenchyme adjacent et le système nerveux,
surtout orthosympathique, jouent un rôle important dans
cette morphogenèse.


