Le système digestif (Cours 3 et 4)
Rappel
 Le système digestif
est constitué d'un tube, formé de plusieurs segments
(bouche - pharynx - oesophage - estomac - intestin grêle
et gros intestin), et de glandes annexes (glandes salivaires,
foie et pancréas). Presque tous les organes digestifs dérivent
de l'endoderme, seuls la bouche et l'anus sont d'origine
mixte: endo et ectodermique.
Le système digestif
est constitué d'un tube, formé de plusieurs segments
(bouche - pharynx - oesophage - estomac - intestin grêle
et gros intestin), et de glandes annexes (glandes salivaires,
foie et pancréas). Presque tous les organes digestifs dérivent
de l'endoderme, seuls la bouche et l'anus sont d'origine
mixte: endo et ectodermique.
La bouche est constituée d'une cavité et d'annexes
spécialisées, les dents, la langue, les glandes
salivaires et les amygdales qui sont des organes lympho-épithéliaux.
Elle est suivie par le pharynx et puis par l'oesophage, première
partie du tube digestif proprement-dit.
La langue est une masse musculaire squelettique recouverte d'une
enveloppe appelée muqueuse qui, dans ce cas, est formée
d'un épithélium et de son chorion. Elle recouvre
le plancher buccal. Son revêtement épithélial
est modifié par des saillies spécialisées:
les papilles linguales.
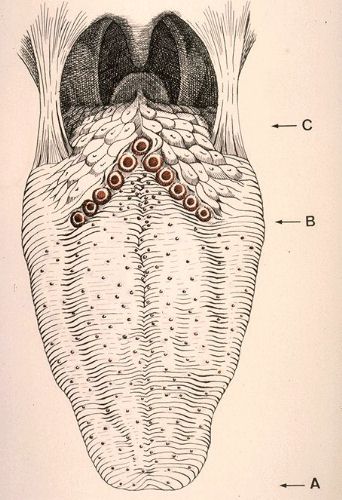
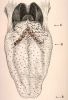 La langue est divisée
en deux parties par le V lingual. La partie antérieure
(A), ou pointe, est mobile et rattachée au plancher
buccal par le frein. Sa face supérieure est parcourue par
un sillon appelé "raphé median".
De part et d'autre du raphé, la surface de la langue est
parcourue de stries transversales et irrégulières
dont l'importance varie d'un individu à un autre. Elles
sont dues à l'alignement des papilles filiformes.
Entre ces stries, se situent de petites saillies très rapprochées,
les papilles fongiformes. Le V lingual (B) dont le sommet
est postérieur, est dû à la disposition des
grandes papilles circonvallariées au nombre de 6
à 12 dans l'espèce humaine. La partie postérieure
(C) ou base de la langue est bosselée parce que la
muqueuse contient en arrière du V lingual, des amas lymphoïdes
dont l'ensemble forme l'amygdale linguale.
La langue est divisée
en deux parties par le V lingual. La partie antérieure
(A), ou pointe, est mobile et rattachée au plancher
buccal par le frein. Sa face supérieure est parcourue par
un sillon appelé "raphé median".
De part et d'autre du raphé, la surface de la langue est
parcourue de stries transversales et irrégulières
dont l'importance varie d'un individu à un autre. Elles
sont dues à l'alignement des papilles filiformes.
Entre ces stries, se situent de petites saillies très rapprochées,
les papilles fongiformes. Le V lingual (B) dont le sommet
est postérieur, est dû à la disposition des
grandes papilles circonvallariées au nombre de 6
à 12 dans l'espèce humaine. La partie postérieure
(C) ou base de la langue est bosselée parce que la
muqueuse contient en arrière du V lingual, des amas lymphoïdes
dont l'ensemble forme l'amygdale linguale.
La couche périphérique
de la langue est un épithélium pavimenteux de type
épidermoïde. Sur la face supérieure,
il est épais et modifié par les papilles. Sur les
bords de la langue (2) et à sa face inférieure,
il est relativement mince. Au niveau du frein (3), il s'infléchit
et devient l'épithélium du plancher buccal. Cet
épithélium repose sur un chorion dense.
La masse de la langue est faite de faisceaux musculaires squelettiques
orientés dans tous les sens et séparés par
des septa conjonctifs où passent les vaisseaux et les nerfs
linguaux. Près du bord de la langue, la masse musculaire
est aussi infiltrée de petites glandes salivaires.
Les modifications épithéliales, très particulières
à la langue, sont des saillies appelées papilles.
Elles sont filiformes, fongiformes ou circonvallariées.
Les papilles filiformes sont les plus fréquentes
et ont un diamètre compris entre 0,3 et 0,5 mm. Dans cette
photographie à la loupe d'une langue humaine,
les papilles filiformes sont toutes des petites saillies en forme
de pointes. Dans d'autres espèces, comme le chat,
leur nombre particulièrement important et leur extrémité
plus effilée donne à la langue le relief d'une râpe.
Chaque papille filiforme
est implantée dans un socle épithélial épais.
Sa paroi est composée de cellules aplaties et fortement
kératinisées dont le grand axe est perpendiculaire
à la surface épithéliale. Le centre de la
papille est occupé par des cellules plus grandes disposées
dans tous les sens. Leur cytoplasme est riche en grains de kératohyaline,
visibles à plus fort grossissement. Seule la partie supérieure
de la papille est creuse. C'est là que desquament les lamelles
kératinisées qui tombent dans la cavité buccale.
Les papilles fongiformes ont un diamètre compris
entre 0,7 et 1 mm. Elles sont dispersées entre les papilles
filiformes et plus nombreuses devant le V lingual. Dans cette
photographie d'une langue humaine,
les papilles fongiformes (flèche) sont des saillies
bombées entre les papilles filiformes. Quelques papilles
fongiformes dans la partie postérieure de la pointe ont
un volume plus important et sont parfois appelées papilles
foliées. Les papilles fongiformes peuvent contenir
des bourgeons gustatifs comparables à ceux que nous
décrirons dans la papille circonvallariée.
A leur niveau, l'épithélium épidermoïde
est plus mince et fait saillie dans la cavité buccale.
L'axe de la saillie est du tissu conjonctif lâche, donc
différent du chorion que l'on trouve en dehors de ces papilles.
La vascularisation importante est responsable de leur teinte rouge
qui ne se perçoit que si l'épithélium desquame
normalement. Dans le cas contraire, les papilles fongiformes sont
masquées et la langue est blanche ou saburrale.
Les papilles circonvallariées
du V lingual sont plus volumineuses que les autres et ont un diamètre
de 1 à 3 mm. Elles forment une saillie arrondie
(flèche) entourée d'un sillon circulaire. Elles
contiennent les organes spécialisés du goût.
Une coupe passant
par le centre de cette papille permet de détailler sa structure
relativement complexe. Le sillon périphérique est
une invagination épithéliale au fond de laquelle
débouchent les petites glandes acineuses séreuses
de Von Ebner. La papille ellemême est une saillie
mamelonnée dont la masse conjonctive est enveloppée
d'une gaine épithéliale criblée de petits
organes en forme de tonnelet, les bourgeons gustatifs.
Le chorion de la papille est lâche et richement vascularisé.
De nombreuses fibres nerveuses amyélinisées
se détachent de la base des bourgeons gustatifs, s'agencent
en petits faisceaux et font synapse avec les volumineuses cellules
nerveuses d'un petit ganglion autonomique situé à
la base de la papille. Les axones des cellules ganglionnaires
se dirigent vers le nerf lingual et le nerf glossopharyngien.
Le bourgeon gustatif
est un organe neuroépithélial dû à
l'invasion de l'épithélium de la papille par des
cellules nerveuses en provenance des crêtes ganglionnaires.
C'est l'organe du goût. Le bourgeon gustatif est
ovoïde. Sa base est en contact avec le chorion lâche.
Son sommet arrive presque à la surface de l'épithélium;
il contient une petite cavité, le canal gustatif qui communique
avec l'extérieur par le pore gustatif.
Les cellules sont allongées et disposées comme les
douves d'un tonnelet. A ce faible grossissement, le pôle
inférieur du bourgeon est flou; on distingue difficilement
les différents types cellulaires et les terminaisons nerveuses
ne sont pas mises en évidence. Les bourgeons sont tassés
les uns contre les autres. Entre eux, l'épithélium
est modifié: les cellules épithéliales sont
perpendiculaires à la surface.
Cette coupe tangentielle
dans la paroi d'une papille circonvallariée démontre
la concentration importante des bourgeons gustatifs à ce
niveau.
L'observation en microscopie électronique
révèle l'existence de quatre types cellulaires
dans le bourgeon gustatif. Ce sont les cellules sombres ou
cellules de type I, les cellules claires ou cellules de
type II, les cellules de type III et les cellules
souches. On ne peut cependant leur attribuer à chacun
un rôle précis. Le canal gustatif est rempli d'une
substance dense et homogène, il reçoit les prolongements
apicaux des différentes cellules.
La cellule de type I allongée possède un
noyau étroit riche en hétérochromatine. Une
expansion du pôle apical munie de longues microvillosités,
se projette dans le canal gustatif. Son cytoplasme contient de
nombreux granules denses, homogènes et limités par
une membrane, qui sont les précurseurs de la substance
dense du canal gustatif. La fonction de cette substance n'est
pas connue.
La cellule de type II possède un grand noyau ovoïde
dont la chromatine est diffuse. Son cytoplasme est moins dense
que celui de la cellule de type I. Il renferme un grand nombre
de vésicules claires. Le pôle apical possède
de courtes microvillosités.
La cellule de type III ressemble à la précédente
par son allure générale et par l'aspect de son cytoplasme.
Son noyau plus irrégulier est plus riche en hétérochromatine.
Les vésicules observées principalement sous le noyau
sont de deux types: les unes, peu nombreuses, ont un contenu dense
et granulaire, les autres plus petites et plus nombreuses ressemblent
fort à des vésicules synaptiques. Cette cellule
est en contact étroit avec des terminaisons nerveuses,
dont le contour est représenté ici en pointillé.
On observe parfois aux zones de contact, un épaississement
de la membrane plasmique et une concentration de petites vésicules.
Les cellules de type I et II sont aussi en contact avec d'autres
terminaisons nerveuses, dont le contour est représenté
ici en trait plein, mais leurs contacts semblent moins spécialisés
car à leur endroit, il n'y a aucun épaississement
de membrane.
Enfin, autour du bourgeon sont appliquées des cellules
épithéliales particulières : les cellules
du type IV ou cellules souches des trois autres types. Elles
migrent dans le bourgeon et s'y différencient.
 Tous les segments du tube digestif
sont formés de quatre couches : la muqueuse, la sousmuqueuse,
la musculeuse et la séreuse. La muqueuse (1)
est la couche la plus interne, qui borde la lumière. Elle
est composée d'un épithélium, différent
d'un segment à l'autre, d'un chorion et d'une bande musculaire
externe : la muscularis mucosae.
Tous les segments du tube digestif
sont formés de quatre couches : la muqueuse, la sousmuqueuse,
la musculeuse et la séreuse. La muqueuse (1)
est la couche la plus interne, qui borde la lumière. Elle
est composée d'un épithélium, différent
d'un segment à l'autre, d'un chorion et d'une bande musculaire
externe : la muscularis mucosae.
La sousmuqueuse (2) est la couche moyenne. Formée
de tissu conjonctif dense, elle contient un réseau vasculaire
important et un ensemble d'éléments nerveux appelé
plexus de Meissner. La musculeuse (3) est la couche externe.
Ces faisceaux musculaires sont généralement concentriques
à la lumière dans la bande interne et disposés
longitudinalement dans la bande externe. Entre les deux bandes,
se trouve un deuxième réseau d'éléments
nerveux appelé plexus d'Auerbach. La séreuse
péritonéale (4) enveloppe la musculeuse de l'estomac
au rectum. Elle est formée d'un mésothélium
et de son chorion riche en cellules adipeuses.
La lumière du tube digestif
est découpée par des évaginations de la paroi:
les valvules conniventes et les villosités.
La valvule connivente se retrouve tout le long du tube digestif.
C'est une évagination de la sousmuqueuse. Elle est
donc formée d'un axe conjonctif qui soulève les
trois couches de la muqueuse. La villosité est spécifique
de l'intestin grêle. C'est une saillie du chorion de la
muqueuse qui soulève l'épithélium.
Voici dans une coupe d'intestin grêle,
une valvule connivente. Son axe conjonctif projette dans la lumière
intestinale les trois couches de la muqueuse. Sa surface
est hérissée de replis plus petits, les villosités
intestinales. Ce sont des expansions épithéliales
dont l'axe conjonctif dépend du chorion de la muqueuse.
Le tissu conjonctif
contient des réseaux nerveux formés de petits nerfs
et de ganglions. Ce sont les plexus de Meissner dans la
sousmuqueuse et d'Auerbach dans la musculeuse. Chaque
nerf, limité par son épinèvre et son périnèvre,
est formé de complexes axoschwanniens entourés
de leur endonèvre. La cellule ganglionnaire est
grande et possède un gros noyau clair excentrique dont
le nucléole est volumineux. Elle est entourée de
cellules gliales, ces cellules capsulaires, dont on ne
voit que le petit noyau foncé.
 L'oesophage est le premier segment du tube digestif proprement-dit.
Examinons sa paroi dans une coupe transversale
photographiée à faible grossissement et à
partir de la cavité centrale située à droite
de l'image. Elle comprend une muqueuse formée d'un épithélium
pluristratifié de type épidermoïde, d'un chorion
dense et d'une muscularis mucosae discontinue. Une épaisse
couche de tissu conjonctif dense, la sousmuqueuse,
sépare la muqueuse de la musculeuse. Celleci
est épaisse et formée de deux couches. L'interne
est circulaire et donc coupée longitudinalement et l'externe
est longitudinale et donc coupée transversalement. Ces
deux couches sont séparées par une fine bande de
tissu conjonctif dense.
L'oesophage est le premier segment du tube digestif proprement-dit.
Examinons sa paroi dans une coupe transversale
photographiée à faible grossissement et à
partir de la cavité centrale située à droite
de l'image. Elle comprend une muqueuse formée d'un épithélium
pluristratifié de type épidermoïde, d'un chorion
dense et d'une muscularis mucosae discontinue. Une épaisse
couche de tissu conjonctif dense, la sousmuqueuse,
sépare la muqueuse de la musculeuse. Celleci
est épaisse et formée de deux couches. L'interne
est circulaire et donc coupée longitudinalement et l'externe
est longitudinale et donc coupée transversalement. Ces
deux couches sont séparées par une fine bande de
tissu conjonctif dense.
L'oesophage ne possède pas de séreuse. Elle est
remplacée par une couche de tissu conjonctif lâche,
l'adventice qui l'unit aux structures voisines.
La muqueuse est
constituée d'un épithélium pluristratifié
de type épidermoïde qui possède des cellules
nucléées même dans ses couches les plus superficielles.
Le chorion formé de tissu conjonctif dense est dépourvu
de glandes. On y observe des fibroblastes, des artérioles
et des veinules. La muscularis mucosae est discontinue.
Ses petits faisceaux de fibres musculaires lisses sont disposés
longitudinalement par rapport à l'axe du tube et donc coupes
transversalement. Ils sont séparés par des travées
de tissu conjonctif dense.
La sousmuqueuse
est une épaisse couche de tissu conjonctif dense
en dehors de la muscularis mucosae. Elle contient
des fibres collagènes et un réseau important de
vaisseaux sanguins.
Dans certaines espèces, la sousmuqueuse contient
des glandes acineuses.
Chez l'homme, ces acini sont muqueux, leurs canaux excréteurs
aboutissent à l'épithélium. Le plexus nerveux
de Meissner n'est pas fort développé vu l'absence
de glandes dans la muqueuse.
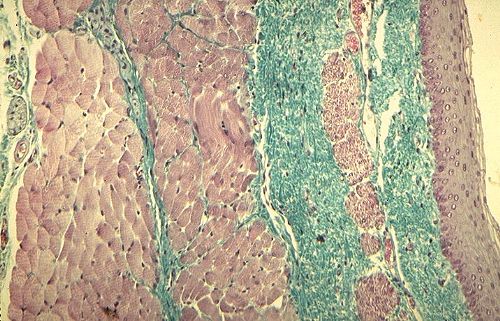
La musculeuse est différente dans la partie supérieure
et inférieure de l'oesophage. Voici sa partie supérieure.
La musculeuse y est formée de deux couches mal délimitées
en de nombreux endroits. Les fibres musculaires y sont surtout
squelettiques car c'est là que se terminent les
muscles constricteurs du pharynx. Dans la partie interne de la
musculeuse, on peut observer de petits faisceaux de muscles lisses
entre les fibres musculaires squelettiques. Ils sont indiqués
par une flèche. Entre les deux couches, se trouvent des
éléments du plexus d'Auerbach disséminés
dans du tissu conjonctif.
Dans la partie inférieure
de l'oesophage, la séparation de la musculeuse en deux
couches est plus nette. La couche interne, circulaire, est un
mélange de fibres musculaires lisses et squelettiques.
Les fibres lisses y prédominent. La couche externe
longitudinale est un mélange à parts égales
des deux types de fibres; les fibres musculaires squelettiques
sont plus colorées. Dans le tissu conjonctif, séparant
les deux couches musculaires, se trouve le plexus d'Auerbach.
Résumons brièvement la structure de la paroi
oesophagienne et la manière de la différencier de
celle des autres segments: sa muqueuse comprend un épithélium
pavimenteux stratifié de type épidermoïde,
un chorion dépourvu de glandes et une muscularis mucosae
discontinue. Sa sousmuqueuse est parfois infiltrée
d'acini muqueux. Sa musculeuse est faite d'un mélange de
muscles lisses et striés.
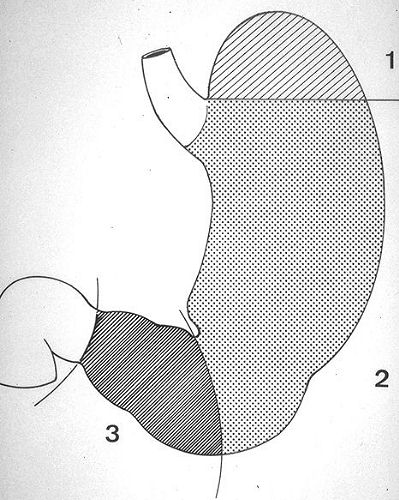
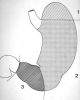 L'estomac est une
vaste poche comprise entre l'oesophage et l'intestin grêle
et située dans l'hypochondre gauche. Il communique avec
l'oesophage par le cardia et avec l'intestin par le pylore. Le
fond ou poche à air (1) est la partie de l'estomac
située audessus d'une ligne horizontale passant par
la limite supérieure du cardia. Le corps de l'estomac
(2) est situé endessous du fond et représente
les 2/3 de son volume. L'antre pylorique (3) conduit au
pylore par le canal pylorique.
L'estomac est une
vaste poche comprise entre l'oesophage et l'intestin grêle
et située dans l'hypochondre gauche. Il communique avec
l'oesophage par le cardia et avec l'intestin par le pylore. Le
fond ou poche à air (1) est la partie de l'estomac
située audessus d'une ligne horizontale passant par
la limite supérieure du cardia. Le corps de l'estomac
(2) est situé endessous du fond et représente
les 2/3 de son volume. L'antre pylorique (3) conduit au
pylore par le canal pylorique.
Lorsque l'estomac est vide, sa surface interne
rose, est hérissée de gros plis longitudinaux ou
obliques anastomosés entre eux. On y observe également
de petites protubérances percées d'un orifice étroit
et circulaire. Ces orifices, les favéoles gastriques,
correspondent en réalité à des cryptes au
fond desquelles débouchent les glandes gastriques.
Nous retrouvons dans la paroi gastrique
la structure générale du tube digestif. La muqueuse
est composée d'un épithélium de revêtement
cylindrique, d'un chorion lâche riche en glandes
et d'une épaisse muscularis mucosae. La sousmuqueuse
est une bande de tissu conjonctif dense richement vascularisé.
La musculeuse est formée de deux couches dont l'interne
et l'externe sont incomplètes. Ces couches sont séparées
par du tissu conjonctif infiltré d'éléments
nerveux. La séreuse entoure l'estomac sauf dans sa partie
supérieure en rapport avec le diaphragme.
 L'épithélium de la muqueuse
est un feuillet glandulaire formé de hautes cellules
cylindriques polarisées. Le noyau est allongé. Une
grosse goutte de mucigène occupe le pôle apical
de toutes les cellules. Le mucus produit est résistant
à l'acide chlorhydrique et à la pepsine et protège
la muqueuse contre sa propre production. Le feuillet glandulaire
s'enfonce dans le chorion au niveau des favéoles gastriques.
L'épithélium de la muqueuse
est un feuillet glandulaire formé de hautes cellules
cylindriques polarisées. Le noyau est allongé. Une
grosse goutte de mucigène occupe le pôle apical
de toutes les cellules. Le mucus produit est résistant
à l'acide chlorhydrique et à la pepsine et protège
la muqueuse contre sa propre production. Le feuillet glandulaire
s'enfonce dans le chorion au niveau des favéoles gastriques.
Par la technique du P.A.S.
on met les glycosaminoglycans et donc le mucus en évidence.
Le chorion est
épais car il contient de nombreuses glandes gastriques.
Leurs coupes sont orientées dans tous les sens suite aux
nombreux replis de la paroi. Le tissu conjonctif qui les entoure
est très lâche et bien vascularisé tout particulièrement
près du feuillet glandulaire. Il contient quelques fibres
musculaires lisses en provenance de la muscularis mucosae.
La partie la plus externe du chorion est une bande de tissu conjonctif
dense appliquée contre la muscularis mucosae. Celleci
est épaisse et continue. On lui distingue parfois deux
ou trois couches de muscles lisses.
La glande gastrique
est une glande tubuleuse aux parois bosselées. Cet
aspect bosselé est particulièrement net dans
cette préparation ou les glandes ont été
dissociées les unes des autres à la suite d'une
perfusion qui a considérablement hydraté
le tissu conjonctif. La lumière glandulaire très
étroite est limitée par deux types cellulaires :
la cellule bordante et la cellule principale. Les
cellules bordantes sont les plus volumineuses et situées
en périphérie. Leur cytoplasme est acidophile. Entre
elles se trouvent les cellules principales, plus petites.
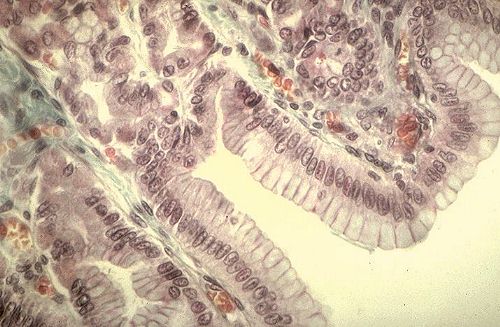
On distingue aux glandes gastriques, trois parties:
le collet, le corps et le fond. Le collet (1) est une région
de transition entre le feuillet glandulaire et le corps de la
glande. Cette région contient des cellules à mucus,
différentes des cellules du feuillet glandulaire et des
cellules bordantes. Dans le corps (2) de la glande, les
cellules à mucus sont presque toutes remplacées
par des cellules principales. La glande se termine par un culdesac
appelé fond (3). Il y a dans la glande gastrique
quatre types cellulaires distincts : la cellule à mucus,
la cellule bordante, la cellule principale et surtout
dans la région pylorique, la cellule endocrine argentaffine
isolée.
La cellule à mucus,
logée entre les cellules bordantes est essentiellement
la cellule du collet. Elle est petite et irrégulière.
Son noyau clair est appliqué au pôle basal et sa
région apicale contient du mucigène P.A.S. positif.
Le mucus produit est différent du mucus libéré
par le feuillet glandulaire. Toutefois, le rôle de cette
sécrétion n'est pas connu.
La cellule bordante
est volumineuse et périphérique. Elle sécrète
l'acide chlorhydrique. Elle peut être sphérique
ou pyramidale et contient un gros noyau. Ses mitochondries sont
particulièrement abondantes mais on n'observe aucun granule
de sécrétion. La membrane du pôle apical de
la cellule bordante, en s'invaginant, une ou plusieurs fois dans
le cytoplasme, forme des replis qui s'ouvrent dans la lumière
de la glande gastrique directement ou par un prolongement qui
se faufile entre les cellules voisines. Ces replis sont de véritables
canalicules dont la paroi possède de nombreuses
microvillosités plus ou moins développées
selon l'état physiologique de la cellule. Le reticulum
endoplasmique rugueux et les ribosomes libres sont peu importants.
L'appareil de Golgi est souvent localisé entre le noyau
et la base de la cellule. Les mitochondries sont réparties
dans tout le cytoplasme. Le réticulum endoplasmique lisse
est formé de vésicules et de tubules courts appelés
généralement "éléments tubulovésiculaires".
Ils sont concentrés autour des canalicules intracellulaires.
Le schéma suivant
montre leurs modifications en fonction de la physiologie cellulaire.
Lorsque la sécrétion d'acide chlorhydrique est stimulée
comme dans le schéma A, les microvillosités s'allongent
et le nombre des éléments tubulovésiculaires
diminue. On admet que tubules et vésicules migrent vers
les canalicules, fusionnent avec eux et accroissent par conséquent
leur étendue en formant les microvillosités.
Lorsque la stimulation est arrêtée (schéma
B), le phénomène inverse se produit : les éléments
tubulovésiculaires réapparaissent en grand
nombre à distance des canalicules dont les microvillosités
ont presque disparu; leur lumière à ce moment paraît
plus large.
Les cellules bordantes sont sécrétrices malgré
leur caractère acidophile. Elles produisent l'acide chlorhydrique
et une glycoprotéine appelée le facteur intrinsèque
qui se lie à la vitamine B12 et facilite son absorption
intestinale.
Voici schématiquement les grandes étapes de la sécrétion d'acide chlorhydrique.
L'acide chlorhydrique n'est pas produit à l'intérieur
du cytoplasme mais dans la lumière des canalicules
où le pH est par conséquent très bas et nettement
inférieur au pH cellulaire. L'acide est le produit de l'attraction
électrostatique des ions chlorures et des protons tous
deux transportés activement à travers la paroi des
canalicules. Les ions chlorures proviennent du sang, les protons
sont libérés indirectement. Des molécules
d'eau réagissent avec l'anhydride carbonique en provenance
du sang pour former de l'acide carbonique sous l'action de l'anhydrase
carbonique. L'acide carbonique s'ionise en proton et en ion bicarbonate.
Le proton est transporté activement vers le canalicule
tandis que l'ion bicarbonate diffuse vers le sang.
La sécrétion d'acide chlorhydrique dépend
de l'anhydrase carbonique car l'inhibition de cet enzyme par l'acetazolamide
empêche sa production.
Les cellules principales
n'existent que dans la moitié inférieure
des glandes gastriques. Elles sont entourées par les volumineuses
cellules bordantes. Le noyau est basal et le cytoplasme, riche
en réticulum endoplasmique rugueux, contient de nombreuses
et grosses vésicules de condensation, peu denses aux électrons.
Leur schéma
est semblable à celui de toute cellule sécrétrice
de protéines. Le pôle apical, hérissé
de petites microvillosités irrégulières,
renferme les vésicules de zymogène rondes
ou ovales. L'appareil de Golgi, bien développé,
occupe la région supranucléaire. Le réticulum
endoplasmique rugueux est important; ses grosses citernes sont
surtout situées à la base de la cellule.
Les granules de zymogène contiennent le pepsinogène,
précurseur de la pepsine. La forme active de l'enzyme n'apparaît
dans la lumière de la glande qu'à pH bas, donc après
production d'acide chlorhydrique et opère de façon
optimale à pH 2. La pepsine est une endopeptidase
qui rompt les liaisons peptidiques dans lesquelles le groupement
aminé appartient à un acide aminé à
cycle aromatique. La cellule principale sécrète
aussi la rennine, enzyme qui précipite la caséine
du lait. Elle forme ainsi du lait caillé dont les protéines
sont soumises ensuite à l'action de la pepsine. Elle est
particulièrement abondante dans le suc gastrique des nourrissons
qui contient moins d'acide chlorhydrique. Elle l'est beaucoup
moins chez l'adulte, chez qui l'acide chlorhydrique est suffisant
pour remplir cette fonction.
Ces glandes typiquement gastriques sont logées dans le
chorion du fond et du corps de l'estomac. Des glandes différentes
existent au niveau du cardia et du pylore.
Les glandes cardiales
sont limitées à une région annulaire étroite
autour de l'embouchure de l'oesophage. Elles sont tubuleuses,
ramifiées et légèrement tordues sur ellesmêmes.
Elles sécrètent du mucus et une petite quantité
d'électrolytes.
Les glandes de l'antre pylorique
sont représentées ici en coupe transversale. Chez
l'homme, elles occupent une région longue d'environ 5 cm.
Dans d'autres espèces, la région des glandes pyloriques
peut occuper jusqu'à 20% de la muqueuse gastrique. Tubuleuses
simples ou ramifiées, elles débouchent dans
des favéoles plus profondes qu'ailleurs. Leurs cellules
sont muqueuses ou endocrines; les cellules pariétales et
bordantes y sont très rares.
 Le tissu conjonctif de la sousmuqueuse
est dense et très vascularisé. Cette couche est
dépourvue de glandes. On y retrouve quelques éléments
nerveux du plexus de Meissner: petits nerfs et cellules ganglionnaires.
Le tissu conjonctif de la sousmuqueuse
est dense et très vascularisé. Cette couche est
dépourvue de glandes. On y retrouve quelques éléments
nerveux du plexus de Meissner: petits nerfs et cellules ganglionnaires.
La musculeuse est
très épaisse et formée de trois couches musculaires
lisses chez l'homme. L'interne est oblique, la centrale est
circulaire et l'externe est longitudinale. Cette dernière
est épaisse au niveau de la petite courbure de l'estomac.
Seule la couche centrale est complète et recouvre toute
la paroi. L'aspect de la musculeuse varie donc d'une région
à l'autre et il est rare d'observer les trois couches dans
une seule coupe. Le tissu conjonctif entre les couches est bien
vascularisé et contient le plexus d'Auerbach. La couche
musculaire externe est recouverte par la séreuse péritonéale.
Les caractéristiques de la paroi gastrique sont les
suivantes.
- La muqueuse est formée d'un feuillet glandulaire,
d'un chorion riche en glandes tubuleuses simples et d'une muscularis
mucosae continue.
- La sousmuqueuse est dépourvue de glandes.
- La musculeuse contient uniquement des muscles lisses
et la séreuse n'est absente que dans la partie supérieure
de l'organe.
La leçon suivante sera consacrée
à l'intestin.
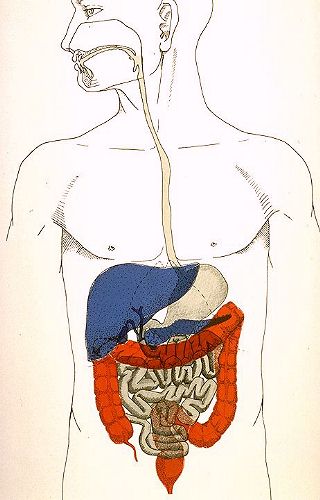 Le système digestif
est constitué d'un tube, formé de plusieurs segments
(bouche - pharynx - oesophage - estomac - intestin grêle
et gros intestin), et de glandes annexes (glandes salivaires,
foie et pancréas). Presque tous les organes digestifs dérivent
de l'endoderme, seuls la bouche et l'anus sont d'origine
mixte: endo et ectodermique.
Le système digestif
est constitué d'un tube, formé de plusieurs segments
(bouche - pharynx - oesophage - estomac - intestin grêle
et gros intestin), et de glandes annexes (glandes salivaires,
foie et pancréas). Presque tous les organes digestifs dérivent
de l'endoderme, seuls la bouche et l'anus sont d'origine
mixte: endo et ectodermique.