Le système digestif (Cours 1 et 2)
 Le système digestif
est un ensemble d'organes dérivant presque tous de l'endoderme.
Seuls la bouche et l'anus sont d'origine mixte: endo- et ectodermique.
D'un point de vue anatomique, la cavité buccale est la
première partie du système. Elle est suivie du tube
digestif proprement dit composé d'une série de segments.
Plusieurs glandes annexes déversent leur produit dans la
lumière de ce tube.
Le système digestif
est un ensemble d'organes dérivant presque tous de l'endoderme.
Seuls la bouche et l'anus sont d'origine mixte: endo- et ectodermique.
D'un point de vue anatomique, la cavité buccale est la
première partie du système. Elle est suivie du tube
digestif proprement dit composé d'une série de segments.
Plusieurs glandes annexes déversent leur produit dans la
lumière de ce tube.
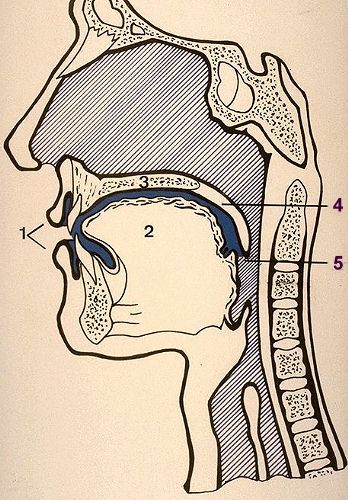
 La cavité buccale
représentée ici en coupe sagittale possède:
La cavité buccale
représentée ici en coupe sagittale possède:
- une paroi antérieure (1) constituée par
les lèvres; deux parois latérales formées
par les joues non visibles sur ce schéma;
- une paroi inférieure composée essentiellement
de la langue (2) et sous la pointe de celleci, d'une petite
région, le "plancher buccal";
- une paroi supérieure constituée par la
voûte palatine (3) et le voile du palais (4);
- et enfin, un orifice irrégulier postérieur,
l'isthme du gosier (5) par lequel elle communique avec le pharynx.
La cavité buccale
est divisée en deux parties inégales par les arcades
dentaires. La partie externe (1) ou vestibule buccal est
la fente située entre les arcades dentaires d'une part,
les joues et les lèvres d'autre part. La partie centrale
(2) est la cavité buccale proprement dite.
Toute la cavité buccale est tapissée d'un épithélium
pavimenteux stratifié de type épidermoïde,
kératinisé seulement au niveau de la langue. Cet
épithélium repose sur un chorion dense dont l'importance
varie d'une région à l'autre. Ce chorion est infiltré
de petites glandes salivaires sauf au niveau des gencives. Ensemble,
l'épithélium et son chorion, forment la muqueuse
buccale.
 Voici cette muqueuse
au niveau du palais. L'épithélium est pavimenteux
stratifié de type épidermoïde et repose sur
un chorion dense très vascularisé. Il est habituellement
infiltré de petites glandes salivaires appelées
"glandes palatines"; elles n'existent pas dans
cette préparation. Ce chorion se confond en profondeur
avec le périoste de la pièce osseuse qu'il recouvre.
Voici cette muqueuse
au niveau du palais. L'épithélium est pavimenteux
stratifié de type épidermoïde et repose sur
un chorion dense très vascularisé. Il est habituellement
infiltré de petites glandes salivaires appelées
"glandes palatines"; elles n'existent pas dans
cette préparation. Ce chorion se confond en profondeur
avec le périoste de la pièce osseuse qu'il recouvre.
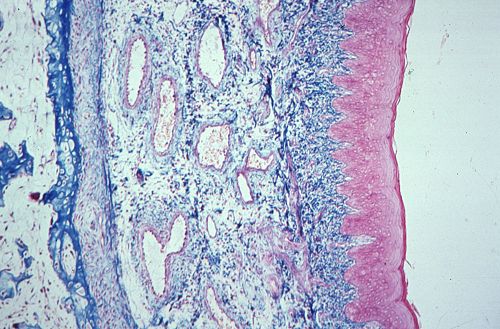
La muqueuse au niveau des parties molles est plus épaisse.
Dans cette coupe de lèvre,
elle double une couche musculaire. Ces fibres musculaires striées
appartiennent aux muscles peauciers. En dehors d'eux, se trouve
la peau formée successivement du derme, riche en glandes
sudoripares et sébacées, puis de l'épiderme.
La région postérieure de la cavité buccale
communique avec le pharynx où se combinent les voies
respiratoires et digestives. Il est subdivisé en trois
parties : la partie supérieure est appelée nasopharynx
(1), la partie moyenne est l'oropharynx (2) et la partie
inférieure, le laryngopharynx (3). L'épithélium
du nasopharynx est respiratoire, donc cylindrique pseudostratifié
alors qu'ailleurs, l'épithélium est épidermoïde
comme dans la cavité buccale et l'oesophage. Le chorion,
très dense, forme l'aponévrose pharyngienne. Son
épaisseur et la distribution des glandes mixtes ou séreuses
qui l'infiltrent varient suivant les endroits. Il est doublé
de nombreux muscles squelettiques.
Les annexes spécialisées de la cavité buccale
sont: les dents, la langue, les glandes salivaires qui
seront étudiées avec les glandes annexes du système
digestif, et les amygdales qui font partie du système
lymphoïde.
Les dents sont des organes minéralisés de consistance
très dure, implantés dans les maxillaires et servant
à la mastication.
 D'un point de vue anatomique, la dent
comprend trois parties : la couronne qui est la partie
visible, la racine incluse dans une logette osseuse, l'alvéole
dentaire, et le collet qui unit la racine à la couronne.
D'un point de vue anatomique, la dent
comprend trois parties : la couronne qui est la partie
visible, la racine incluse dans une logette osseuse, l'alvéole
dentaire, et le collet qui unit la racine à la couronne.
Les dents sont
classées d'après leur forme et leur situation en
incisives, canines, prémolaires et molaires. Les
incisives sont antérieures, leur couronne est tranchante
et leur racine est simple. Les canines sont latérales,
leur couronne est pointue et leur racine est simple. Les prémolaires
sont latérales, leur couronne est cubique et possède
une face triturante munie de deux tubercules ou cuspides. Leur
racine est généralement simple. Les molaires sont
postérieures, leur couronne est cubique et possède
une face triturante formée de quatre cuspides. Leurs racines
sont doubles ou triples. Dans cette diapositive, les dents sont
vues par leur face interne.
Le nombre de dents se calcule par demi-face et s'exprime par la
formule dentaire. Elle est différente dans la dentition
de lait et dans la dentition définitive. Dans la dentition
de lait, il y a dans chaque demi-mâchoire deux incisives,
une canine et deux prémolaires. Dans la dentition définitive,
il y a dans chaque demi-mâchoire, deux incisives, une canine,
deux prémolaires et trois molaires.
Pendant l'enfance, les deux dentitions coexistent. Cette radiographie
le démontre: certaines dents définitives sont encore
incluses sous les racines des dents de lait. L'éruption
des dents s'étend depuis le quatrième mois jusqu'à
la vingtième année. La première molaire définitive
est en position occlusale à l'âge de six ans.
La dent est creusée
d'une cavité centrale, la cavité pulpaire
(1), qui s'ouvre à l'extrémité de la racine
par l'apex (2). Elle est entourée d'une substance
minéralisée, la dentine (3), couverte au
niveau de la couronne par l'émail (4) et au niveau
de la racine par le cément (5). Le ligament alvéolo-dentaire
(6) formé de faisceaux fibreux joint les parois de l'alvéole
(7) au cément de la racine.
Pour mieux comprendre la structure de la dent, il est nécessaire
de connaître sa genèse. Voici quelles en sont les
différentes étapes.
Au premier mois de la vie embryonnaire,
la cavité buccale primitive (1) est entourée
du deuxième arc branchial, l'arc mandibulaire (2),
d'où sont issus les bourgeons qui forment l'arc maxillaire
(3). Elle est surplombée par la saillie du cerveau primitif
(4).
Les deux arcs sont composés d'une masse mésenchymateuse,
enveloppée d'une lame ectoblastique. Les trois schémas
représentent trois stades précoces de leur évolution.
En A : une coulée ectoblastique s'enfonce dans le mésenchyme
sous-jacent : c'est le mur plongeant.
En B : cette coulée se creuse et la fente ainsi formée
devient en s'élargissant le vestibule buccal qui
sépare la partie externe (futures lèvres ou futures
joues) de la partie interne (futur maxillaire). A partir du mur
plongeant naît une deuxième coulée ectoblastique,
la lame dentaire, qui s'enfonce aussi dans le mésenchyme.
En C : la lame dentaire produit en certains points de petits nodules
épithéliaux, les premiers bourgeons dentaires.
Dès que les bourgeons sont formés, la lame dentaire
dégénère. Seule sa partie interne persiste
pour donner une deuxième série de bourgeons (flèche),
ceux des dents définitives en dedans des précédents.
Le bourgeon épithélial (A) naît de
la lame dentaire
ectoblastique. Il induit la condensation du mésenchyme
voisin en une papille (B). Essentiellement mésoblastique,
cette papille contient quelques cellules ectoblastiques en provenance
des crêtes neurales et représentées en vert
dans le schéma. Entretemps, la lame dentaire produit un
deuxième bourgeon, celui de la dent définitive.
En C, le bourgeon initial se déprime en cloche et devient
l'organe adamantin. La lame dentaire dégénère.
Le tissu mésenchymateux s'organise autour de l'organe adamantin
et de la papille; il devient le sac dentaire.
Ces trois ébauches primitives sont à l'origine
de la dent. L'organe adamantin, ectoblastique, produira l'émail.
La papille, ectomésoblastique, produira la pulpe et la
dentine. Le sac dentaire, mésoblastique, produira le cément
et le ligament alvéolo-dentaire.
L'organe adamantin
est l'inducteur de l'évolution dentaire. Son bord (1) comprend
deux couches cellulaires accolées. Il induira la formation
de la racine. Ailleurs (2), l'organe adamantin, formé de
trois couches cellulaires, est à l'origine de la couronne.
Voici la coupe
qui correspond au schéma de l'image précédente.
L'organe adamantin comprend trois couches. Le mur externe
est une lamelle irrégulière de petites cellules.
Il est en rapport avec le sac dentaire. La couche moyenne est
appelée "reticulum adamantin'' parce qu'elle
est formée de petites cellules étoilées anastomosées
les unes aux autres en un réseau dont les mailles contiennent
une substance riche en mucopolysaccharides. Le mur interne
est une palissade de cellules cylindriques relativement hautes,
les préadamantoblastes, en relation avec la papille mésenchymateuse.
A l'endroit ou les deux murs se rejoignent (flèche), le
réticulum est très peu important et l'organe adamatin
est pratiquement réduit à deux couches. Ces deux
couches accolées forment le feuillet de Hertwig
dont dépendra l'évolution de la racine.
En dedans du bourgeon que nous venons de décrire, se trouve
l'ébauche du bourgeon de la dent définitive.
Le principe général de la formation de la couronne
est le suivant. Là où il existe du réticulum,
les préadamantoblastes induisent d'abord la différenciation
des cellules mésenchymateuses de la papille en odontoblastes.
Ceux-ci produisent ensuite de la dentine. Au contact de
cette première couche de dentine, les préadamantoblastes
se transforment alors en adamantoblastes, cellules spécialisées
dans la sécrétion de l'émail.
Schématisons ces différentes étapes.
Avant toute différenciation (schéma A), les préadamantoblastes
forment dans l'organe adamantin une palissade entre le réticulum
adamantin et la papille mésenchymateuse richement vascularisée.
Très rapidement, (schéma B), ils induisent la différenciation
des cellules mésenchymateuses adjacentes en odontoblastes.
Une fois en place, (schéma C), les odontoblastes sécrètent
la première couche de dentine. Ce premier dépôt
de dentine induit immédiatement la transformation des préadamantoblastes
en adamantoblastes. Ils s'allongent et changent de polarité.
Dès leur différenciation, (schéma D), les
adamantoblastes déposent une couche d'émail
contre la face externe de la dentine. La présence du réticulum
adamantin est indispensable à ce dépôt.
Les odontoblastes
déposent une seconde couche de dentine à la face
interne de la première (schéma 1) et peu après,
les adamantoblastes déposent une seconde couche d'émail
à la face externe de la première (schéma
2). Dès qu'ils sont entamés, ces deux processus
continuent simultanément (schéma 3). Plusieurs couches
d'émail et de dentine se déposent de telle façon
que, par rapport à la papille, l'émail semble
s'accroître de façon centrifuge et la dentine
de façon centripète.
La croissance de l'émail
écrase progressivement les adamantoblastes, le réticulum
adamantin, le mur externe et le sac dentaire. Lorsque la production
d'émail est suffisante, les quatre couches sont comprimées;
elles dégénèrent et forment une lame contenant
de nombreux débris cellulaires, la membrane de Nasmyth
(flèche) éliminée lors de l'éruption.
Puisqu'il n'y a plus d'adamantoblastes, la production d'émail
s'arrête. La dentine, par contre, continue à
être élaborée. Ses couches successives refoulent
les odontoblastes vers le centre de la pulpe qui diminue sans
cesse.
Avant son éruption,
la dent ne possède qu'une racine incomplète. La
formation de la racine définitive dépend du feuillet
de Hertwig (flèche). Celui-ci est la seule partie de
l'organe adamantin qui n'est pas éliminée lors de
l'éruption. Il provient du bord périphérique
de cet organe où mur interne et externe sont accolés.
La formation de la racine
s'effectue en plusieurs étapes.
- Les préadamantoblastes induisent d'abord la formation
d'odontoblastes. Ceux-ci produisent ensuite une couche
de dentine.
- Parce qu'il n'y a pas de réticulum, la production d'émail
est impossible; donc, les cellules du feuillet de Hertwig dégénèrent
et la dentine induit alors la formation de cémentoblastes
à partir des cellules mésenchymateuses du sac dentaire.
- Ceux-ci produisent du cément et se transforment
en cémentocytes.
Le schéma A représente le feuillet de Hertwig. Son
extrémité n'est pas différenciée.
Elle sera le siège de nombreuses mitoses qui entraîneront
sa migration progressive le long de la papille mésenchymateuse
(1). Dans sa partie supérieure, il comprend les deux murs
dont l'interne (2) est une palissade de quelques cellules
équivalentes aux préadamantoblastes décrits
au niveau de la couronne. En dehors du feuillet de Hertwig se
trouve le sac dentaire (3).
Les schémas suivants expliquent comment la racine croît
segment par segment. Dans chaque segment représenté,
la suite des événements est identique. Pendant que
le feuillet de Hertwig s'enfonce (schéma B), son mur interne
induit la formation d'odontoblastes (4) dans la papille.
Ces odontoblastes élaborent une première couche
de dentine (5) (schéma C). Vu l'absence de réticulum
adamantin, les cellules du feuillet qui ont induit la formation
de ces premiers odontoblastes dégénèrent
et le sac dentaire est en rapport immédiat avec la dentine
(schéma D). Les cellules mésenchymateuses du sac
dentaire prennent appui sur la dentine et se transforment en cémentoblastes
(6) (schéma E). Ceux-ci se comportent comme des ostéoblastes
: ils s'entourent de leur propre production, le cément
et deviennent des cémentocytes (schéma F).
Résumé
La formation de la dent dépend donc de l'organe adamantin
dont la première fonction est d'induire la transformation
des cellules mésenchymateuses de la papille en odontoblastes.
Au niveau de la couronne, il produit l'émail puis
disparaît. La couronne est donc élaborée rapidement
et sa taille ne sera plus modifiée après l'éruption
dentaire. Au niveau de la racine, le feuillet de Hertwig
est maintenu. Il induit la formation de la racine dont la croissance
est progressive et ne s'arrêtera qu'au moment où
le pouvoir mitotique du feuillet est épuisé.
Les adamantoblastes sont des cellules d'origine ectoblastique
et produisent l'émail. Ils proviennent de la différenciation
du mur interne de l'organe adamantin.
Dans cette coupe de bourgeon dentaire,
les trois couches de l'organe adamantin : le mur externe, le
réticulum adamantin et le mur interne sont bien visibles.
Les adamantoblastes du mur interne sont disposés régulièrement
et appliqués contre la couche des tissus calcifiés.
En-dedans de ceux-ci se trouvent les odontoblastes et la
pulpe. L'ensemble est entouré par le tissu mésenchymateux
du sac dentaire, très peu organisé à ce stade
du développement.
Les adamantoblastes sont de hautes cellules cylindriques.
Ils sont observés ici en coupe semi-fine,
colorée au bleu de toluidine et agrandie environ 1.200
fois.
Le noyau est situé dans le pôle de la cellule orienté
vers le réticulum adamantin, qui n'est pas visible sur
la photo. Le pôle opposé est incrusté dans
une bande, teinte en bleu foncé et qui représente
les premières couches d'émail calcifié...
Le corps très grêle des adamantoblastes contient
quelques points bleus de taille variable. Ce sont des corps
adamantins. Seuls ceux dont le volume est suffisant, peuvent
être observés à ce grossissement.
Le prolongement de Tomes est la petite partie plus claire
de la cellule appliquée contre l'émail. Son extrémité
est fréquemment insérée dans un espace conique
qui, dans cette coupe, prend l'aspect d'une petite incisure en
forme de V.
Les adamantoblastes
sont unis entre eux par des complexes de jonction situés
près de leurs deux pôles. Les complexes de jonction
apicaux marquent la limite entre le corps cellulaire et le prolongement
de Tomes. L'adamantoblaste, qui produit l'émail, possède
les caractères d'une cellule sécrétrice:
un réticulum endoplasmique rugueux important et un appareil
de Golgi bien développé. Les mitochondries sont
nombreuses au pôle basal de la cellule. Des granules denses
limités par une membrane sont parsemés dans le cytoplasme
et accumulés dans le prolongement de Tomes : ce sont les
corps adamantins.
Cette suite de schémas
explique la sécrétion des corps adamantins.
Certains sont excrétés au niveau des faces latérales
de la cellule. Ils seraient à l'origine de la substance
interprismatique de l'émail. La plupart sont accumulés
dans le prolongement de Tomes (schéma 1). L'excrétion
en bloc de ces corps adamantins accumulés (schéma
2) produit un segment de prisme et la cellule semble décapitée.
Mais elle reconstitue un prolongement (schéma 3) où
s'accumule un nouvel amas de corps adamantins dont l'excrétion
(schéma 4) produit un nouveau segment de prisme. La substance
interprismatique et les prismes seront expliqués en décrivant
la structure de l'émail.
L'émail est le tissu calcifié le plus dur de
l'organisme. Il est d'origine ectoblastique et recouvre la dentine
au niveau de la couronne.
Comme tout autre tissu minéralisé, l'émail
possède une phase organique et une phase minérale.
La phase organique ne représente cependant que 3 % de son
volume et est dépourvue de collagène. La phase minérale
est donc particulièrement importante. Elle est purement
cristalline. Ses cristaux d'hydroxyapatite sont plus volumineux
que dans les autres tissus minéralisés. L'émail
contient aussi un pour mille de cristaux de fluoroapatite
responsables de sa grande dureté.
A faible grossissement et sur coupe obtenue par usure,
l'émail paraît strié de lignes perpendiculaires
à la surface de la dent et de bandes moins nettes et plus
ou moins parallèles à cette surface. Les premières
délimitent les unités de l'émail appelées
prismes. Les autres sont les lignes d'accrétion
: elles marquent les dépôts successifs réalisés
de façon synchrone par tous les adamantoblastes.
Le schéma suivant
permet de les comprendre.
Les lignes perpendiculaires, flexueuses, s'étendent de
la dentine à la surface de la dent. Elles délimitent
un fragment d'émail, le prisme (1). La ligne elle-même
est la membrane prismatique (2) et deux prismes sont séparés
par une très étroite bande de substance interprismatique
(3). Chaque prisme est formé par la superposition des petits
segments (4) déposés successivement par un
adamantoblaste. Parce que la production de tous les adamantoblastes
est synchrone, les segments de prismes voisins sont régulièrement
juxtaposés...
Cette juxtaposition détermine les lignes d'accrétion
(5). Certaines sont plus épaisses que d'autres, elles traduisent
un arrêt du dépôt et donc un trouble physiologique.
L'une d'elles, la ligne néonatale, (6) est constante
: elle marque le trouble physiologique profond de la naissance.
Le prisme peut donc se définir de deux façons. C'est
d'une part, l'unité structurelle de l'émail.
Il a la forme d'un bâtonnet sinueux qui s'étend de
la jonction entre l'émail et la dentine jusqu'à
la surface de la dent. Il résulte de la superposition d'une
série de segments.
En coupe transversale,
les prismes peuvent avoir des formes très diverses. La
forme prismatique, d'où les prismes tirent leur nom, est
classique mais plutôt rare. On observe parfois des sections
transversales tout à fait circulaires ou en fer à
cheval.
La subdivision de l'émail en prismes, membranes prismatiques
et substance interprismatique ne repose sur la différenciation
d'aucune structure précise au sens habituel du terme. Elle
est due uniquement à la disposition des cristaux d'hydroxyapatite,
perceptible dans cette micrographie électronique.
Substance prismatique (A) et interprismatique (B) diffèrent
donc essentiellement par l'orientation de leurs cristaux.
La membrane prismatique est plus riche en substance organique
et les cristaux y sont plus rares.
Les odontoblastes sont des cellules mésenchymateuses
issues de la papille dentaire; elles produisent la dentine.
Les odontoblastes
induits par les préadamantoblastes forment la couche
externe de la papille. Le corps cellulaire est cylindrique
et basophile. On peut y observer une tache plus claire due à
l'importance du système de Golgi. Le noyau est situé
au pôle basal, orienté vers la papille. Le pôle
apical est appliqué contre une bande claire, la prédentine.
Il est prolongé par une expansion très grêle,
la fibre de Tomes, qui n'est pas visible dans cette coupe
légèrement oblique. Elle s'insinue plus ou moins
loin dans un petit canalicule dentinien qui s'étend jusqu'à
l'émail.
Ce schéma
explique l'ultrastructure des odontoblastes et démontre
leur fonction sécrétoire. Le réticulum endoplasmique
rugueux est particulièrement important et l'appareil de
Golgi est bien développé. Le cytoplasme contient
de larges vésicules claires et des grains de sécrétion.
Les cellules sont unies entre elles par divers complexes de jonction
le long de leurs faces latérales. Ils sont plus importants
au pôle apical.
La dentine est une substance blanchâtre et dure. Elle
est produite par les odontoblastes. Elle est traversée
par de nombreux canalicules où se trouvent les fibres de
Tomes, seuls éléments cellulaires de ce tissu.
Comme tous les tissus minéralisés, la dentine possède
deux phases: une phase organique et une phase minérale.
La phase organique représente 30 % de son volume
et est composée de fibres collagènes et de substance
fondamentale. La phase minérale est formée
d'une fraction amorphe très importante riche en carbonates
et d'une fraction cristalline constituée de cristaux d'hydroxyapatite.
Examinons la dentine dans une coupe de bourgeon dentaire
colorée selon une méthode trichromique classique.
Elle est située en périphérie de la papille
mésenchymateuse. Sa couche interne, la plus mince, est
colorée en bleu. C'est la couche la plus récente
ou prédentine qui n'est pas minéralisée.
La couche la plus épaisse est la dentine vraie minéralisée.
Les deux couches sont parcourues de lignes parallèles,
les canalicules dentiniens qui contiennent les fibres de
Tomes. Ceux-ci permettent de différencier clairement la
dentine de l'émail qui la recouvre.
Dans une dent adulte,
et sur coupe longitudinale usée et colorée au bleu
de toluidine, la distinction entre prédentine et dentine
vraie n'est plus possible, au moins à ce faible grossissement.
Les canalicules dentiniens sont ces fines lignes sombres
et parallèles qui s'étendent de la cavité
pulpaire à l'émail. Dans d'autres types de préparation,
il est possible d'observer en outre des lignes parallèles
à la surface. Ce sont les lignes d'accrétion
et, comme dans l'émail et pour les mêmes raisons,
la ligne néonatale est plus prononcée.
La dentine peut
être subdivisée en dentines intertubulaire
et péritubulaire.
La dentine intertubulaire est produite en plusieurs étapes.
Les odontoblastes produisent du précollagène et
les précurseurs de la substance fondamentale dont on retrouve
les premières traces dans leurs vésicules et leurs
grains de sécrétion. Les précurseurs déposés
à la surface apicale des cellules forment une couche argentaffine,
la prédentine précoce (schéma A).
Pendant le dépôt d'une deuxième couche, la
première se modifie, elle perd son affinité pour
l'argent et devient la prédentine (schéma
B). Dans une étape ultérieure (schéma C),
des points de calcification ou calcosphérites apparaissent
dans la matière organique. Ce début de calcification
caractérise la dentine intermédiaire. Les calcosphérites
croissent de façon radiaire et fusionnent. La dentine est
ainsi entièrement calcifiée et devient la dentine
vraie (schéma D).
A chaque dépôt d'une nouvelle couche, la fibre
de Tomes s'accroît d'autant. La dentine péritubulaire
n'est produite que dans la dentine intertubulaire calcifiée.
Son mode de production est identique.
La distinction entre dentines intertubulaire et péritubulaire
est nette dans une coupe transversale,
examinée à fort grossissement.
La zone plus claire près de la pulpe est de la prédentine
précoce (1). Elle est longée en-dehors par une
bande de dentine intermédiaire (2). Les deux zones
sont parsemées de petites cavités, les coupes des
canalicules dentiniens. Certaines contiennent un point
bleu, la fibre de Tomes. Dans la dentine intermédiaire,
chaque canalicule est entouré d'un halo plus foncé
que le tissu voisin. Ce halo correspond à la dentine
péritubulaire qui n'est produite que dans les régions
calcifiées de la dentine intertubulaire.
Le rythme et la régularité du dépôt
permettent de distinguer plusieurs variétés de dentine.
La dentine primaire est antérieure à l'éruption
dentaire. Son dépôt est rapide et régulier.
La dentine secondaire est postérieure à l'éruption
dentaire. Son dépôt est lent et régulier.
La dentine tertiaire est une dentine de réparation.
Son dépôt est rapide et irrégulier.
La pulpe est un
tissu mésenchymateux richement vascularisé
et richement innervé. Elle est le reste de la papille
dentaire embryonnaire et remplit la cavité centrale de
la dent. Sa couche la plus externe est celle des odontoblastes
appliqués contre la dentine, reconnaissable à ses
tubules.
Son volume varie avec l'âge: considérable
chez l'enfant, il diminue chez l'adulte et est réduit à
rien chez le vieillard chez qui la pulpe finit même par
disparaître complètement. L'innervation de la pulpe
et de la dentine est mal connue. Une branche nerveuse se détache
du nerf dentaire et pénètre par l'apex dans la pulpe.
Elle y perd sa gaine de myéline et ses fibres se dirigent
vers les odontoblastes autour desquels elles s'anastomosent en
réseau. Quelques rameaux issus de ce réseau s'enroulent
autour du corps d'un odontoblaste puis de sa fibre de Tomes dans
le canalicule dentinien. Les autres restent dans la pulpe et enveloppent
la base des odontoblastes.
La présence de fibres nerveuses dans quelques canalicules
n'est pas suffisante pour expliquer la douleur dentinienne. Celle-ci
est le résultat d'une action mécanique exercée
par les cellules sur le plexus nerveux. Cette action est provoquée
par l'aspiration de la fibre de Tomes dans le canalicule ou par
son expulsion hors du canalicule.
Le cément
est un tissu minéralisé qui enveloppe la
dentine au niveau de la racine. Il est élaboré par
les cémentoblastes qui s'entourent de leur production
et deviennent des cémentocytes. Parmi tous les tissus
calcifiés de la dent, il est donc celui qui ressemble le
plus au tissu osseux.
Il est formé de lamelles parallèles représentées
ici à fort grossissement. Chacune d'elles est creusée
de petites cavités où sont logés les cémentocytes.
Ces logettes sont reliées entre elles par de fins
canalicules.
Le cément (1) est uni à la paroi de la cavité
alvéolaire (2) par des faisceaux fibreux dont l'ensemble
est appelé "ligament alvéolo-dentaire".
Les espaces relativement larges entre faisceaux fibrillaires sont
occupés par des vaisseaux sanguins. Ce ligament assure
le maintien de la dent dans son alvéole.
Nous avons ainsi terminé la description de la dent. La
leçon prochaine sera consacrée
à la langue et au début de l'étude du tube
digestif proprement-dit.
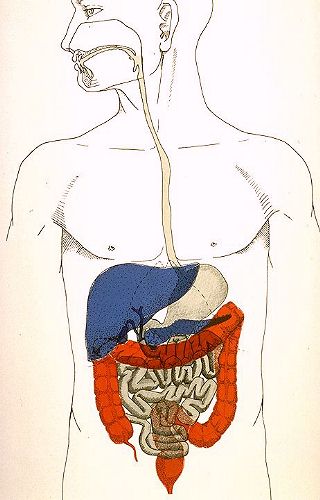 Le système digestif
est un ensemble d'organes dérivant presque tous de l'endoderme.
Seuls la bouche et l'anus sont d'origine mixte: endo- et ectodermique.
D'un point de vue anatomique, la cavité buccale est la
première partie du système. Elle est suivie du tube
digestif proprement dit composé d'une série de segments.
Plusieurs glandes annexes déversent leur produit dans la
lumière de ce tube.
Le système digestif
est un ensemble d'organes dérivant presque tous de l'endoderme.
Seuls la bouche et l'anus sont d'origine mixte: endo- et ectodermique.
D'un point de vue anatomique, la cavité buccale est la
première partie du système. Elle est suivie du tube
digestif proprement dit composé d'une série de segments.
Plusieurs glandes annexes déversent leur produit dans la
lumière de ce tube.