NERVOUS TISSUE
The nervous system consists of all nervous tissue in the body. It is divided anatomically into the central nervous system and the peripheral nervous system.
The CNS consists of the brain (encephalon), which is enclosed in the skull, and the spinal cord, which is contained within the vertebral canal. Nervous tissue of the CNS does not contain connective tissue other than that in the meninges and in the walls of large blood vessels. Collagenous fibers or fibrocytes/blasts are consequently not observed, which is quite unlike other tissues. Because of the absence of connective tissue, fresh CNS tissue has a very soft, somewhat jelly-like consistency. The two major classes of cells that make up the nervous tissue are nerve cells, neurones, and supporting cells,
glia.
The vast majority of neurones is generated before birth. Persisting stem
cells give rise to a small number of new neurones
troughout the lifetime of mammals, including humans. The permanent addition
of neurones may be important for the maintenance and plasticity of some parts
of the CNS, but it is insufficient to replace neurones that die because of
disease or acute damage to the CNS. Neurones are "intended" to last
a lifetime. Mature neurones are not mitotically active, i.e. they do not divide.
Neurones are generally large cells. Neural activity and its control require
the expression of many genes, which is reflected in the large and light nuclei
of most neurones. The keys to the understanding of the function of a neurone
lies in (1) the shape of the neurone and, in particular, its processes, (2)
the chemicals the neurone uses to communicate with other neurones (neurotransmitters)
and (3) the ways in which the neurone may react to the neurotransmitters released
by other neurones.
-
Neurones have long processes (neurites),
which extend from the part of the cell body around the nucleus, the perikaryon
or soma. Neurites can divided into two
functionally and morphologically different groups - dendrites
and axons.
Dendrites are part of the receptive surface
of the neurone. As a rule neurones have one to several primary
dendrites, which emerge from the perikaryon.
Primary dendrites may divide into secondary, tertiary
etc. dendrites.
Each neurone has as a rule one axon, and never
more than one axon which emerges from the perikaryon or close to
the trunks of one of the primary dendrites. The point of origin of the
axon from the perikaryon is the axon hillock.
The axon may, like the dendrites, branch as it travels through the nervous
tissue to its destination(s). The axon is the "transmitting"
process of the neurone.
The shape and orientation of the dendritic tree of the neurone determines
the amount and type of information that may reach the neurone. The course
of its axon determines to which neurones this information may be passed
on. The location of the neurone within the CNS determines to which major
system the neurone belongs.
There are several hundred functionally different areas,
i.e. groups of neurones, in the CNS. Based on location, shape of the dendritic
tree and course of the axon, several thousand types of neurones can be
distinguished in the CNS.
-
Neurotransmitters either excite or inhibit the postsynaptic neurone.
Neurotransmitters are found in vesicles within bulb-shaped swellings of
its axon, synaptic vesicles in synaptic
boutons. The most prominent excitatory transmitter in the CNS is
glutamate. The most prominent inhibitory
transmitter in the CNS is GABA (gamma-amino
butyric acid). Other "main" neurotransmitters are e.g. dopamine, serotonin,
acetylcholine, noradrenaline and glycine. Each neurone uses only one of
the main transmitters, and this transmitter is used at all synaptic boutons
that originate from the neurone.
One or more of the "minor" transmitters (there are
several dozens of them - such as cholecystokinin, endogenous opioids,
somatostatin, substance P) may be used together with a main transmitter.
-
Usually there exists a multitude of receptors which are all sensitive
to one particular neurotransmitter. Different receptors have different
response properties, i.e. they allow the flux of different ions over the
plasma membrane of the neurone or they may address different second messenger
systems in the postsynaptic neurones. The precise reaction of the neurone
to the various neurotransmitters released onto its plasma membrane at
the synapses is determined by the types of receptors expressed by the
neurone. Several
functionally different receptors exist for each of the major neurotransmitters.
- Suitable Slides
- sections of spinal cord
- H&E, luxol fast
blue/cresyl violet (LFB/CV), toluidine blue, Giemsa
Thoracic Spinal Cord, sheep - LFB/CV
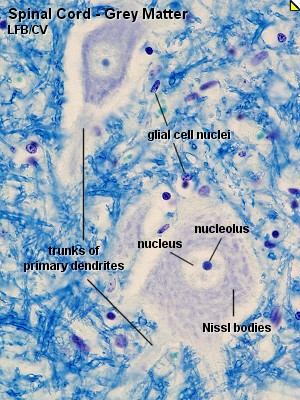
Most neurones have a light, large nucleus with a distinct nucleolus. The cytoplasm
of many neurones contains fairly large amounts of rough endoplasmatic reticulum,
which may aggregate within the cytoplasm of the neurone to form Nissl-bodies.
Nissl-bodies are prominent in motor neurones located in the ventral horn of
the grey matter of the spinal cord. The neurites are difficult to identify
in most types of stained sections. Only the most proximal segments of the
primary dendrites are seen clearly. The size of the perikaryon depends on
the level of activity of the neurone and the length of the processes which
the neurone has to support. An usable range for the size of the perikaryon
would be 15-50 µm, although much smaller and much larger neuronal perikarya
exist.
Draw the spinal cord at low magnification and indicate
the distribution of grey matter and white matter. Find a nice group of neurones
in the grey matter and draw them at a high magnification. Finally, have a
look at the white matter and identify the nuclei of glial cells. You will
find similar nuclei also in the grey matter.

Thoracid spinal Cord - H&E, silver stain
These slides show the same major features as the LFB/CV stained sections.
Try to identify neurones (primary dendrites, Nissl-bodies) and glial cell
nuclei in the H&E stained section. Part of the cytoskeleton of neurones
is (like the reticular connective tissue fibers)
argyrophilic, i.e. they "love" silver and can be stained by silver stains.
Aside from the neurones and their processes, fine fibrils are visible in the
neuropil. Many of the fibrils represent axons travelling in the grey and white
matter of the spinal cord.
It should not be necessary to prepare separate drawings
for these slides. Make sure that you can identify the main structural features
in all preparations.


CNS tissue contains several types of non-neuronal, supporting cells, neuroglia.
- Astrocytes (or astroglia) are star-shaped cells.
Their processes are often in contact with a blood vessel (perivascular
foot processes). Astrocytes provide physical and metabolic support
to the neurones of the CNS. They participate in the maintenance of the composition
of the extracellular fluid. Although not themselves directly
involved in the process of communication between neurones, they may be involved
in the removal of transmitters from synapses and the metabolism of transmitters.
Astrocytes are also the scar-forming cells of the CNS.
- Oligodendrocytes (or oligoglia) have fewer and shorter processes. Oligodendrocytes form myelin in the white matter of the CNS and are the functional homologue of peripheral Schwann cells. Oligodendrocytes may, in contrast to Schwann cells in the periphery, form parts of the myelin sheath around several axons.
- Microglia are small cells with complex shapes. Microglia are, in contrast to neurones and the other types of glial cells, of mesodermal origin. They are derived from the cell line which also gives rise to monocytes, i.e. macrophage precursors which circulate in the blood stream. In the case of tissue damage, microglia differentiate into phagocytotic cells.
- The ventricles of the brain and the central canal of the spinal cord are
lined with ependymal cells. Regionally these
cells may differentiate into specialised tanycytes.
Many glial cells do express neurotransmitter receptors, but they do not form synapses with neurones. Neuronal activity may regulate glial function by a spillover of transmitter from synaptic sites, which are typically surrounded by fine processes of glial cells. Glial cells may also communicate with each other via GAP junctions.
- Suitable Slides
- sections of the forebrain
- toluidine blue, Giemsa, luxol fast blue/cresyl
violet
Forebrain, Cortex, mouse - Giemsa and Forebrain,
Hippocampus, mouse - Giemsa
Most glial cells are much smaller than neurones. Their nuclei are generally
much smaller than neuronal nuclei, and they rarely contain an easily visible
nucleolus. Other aspects of their morphology are variable. The glial cytoplasm
is, if visible at all, very weakly stained. Different types of glial cells
cannot be easily distinguished by their appearance in this type of preparation.
Most of the small nuclei located in the white matter of the CNS, where they
may form short rows, are likely to represent oligodendrocytes.
Browse through the sections at low or medium magnification and try to get
a feeling for the structural diversity visible in the section available to
you - parts of the section that look different from others are very likely
to have different functions.
Find a spot that appears interesting (or
least boring) to you and sketch its structure at low magnification.
Choose a spot for high magnification, and draw some of the visible neurones
and glial cells. Note the difference in the size and number of glial cells
and neurones.


The PNS comprises all nervous tissue outside the brain and spinal cord. It
consists of groups of neurones (ganglion cells),
called ganglia, feltworks of nerve fibres,
called plexuses, and bundles of parallel nerve
fibres that form the nerves and nerve roots.
Nerve fibres, which originate from neurones within the CNS and pass out of
the CNS in cranial and spinal nerves, are called efferent
or motor fibers. Nerve fibres which originate
from nerve cells outside the CNS but enter the CNS by way of the cranial or
spinal nerves are called afferent or sensory
nerve fibres.
The principal neurotransmitters in the PNS are acetylcholine
and noradrenalin.
Afferent, sensory fibres enter the spinal cord via
the dorsal roots, while efferent, motor fibres leave the spinal cord via the
ventral roots. Dorsal and ventral roots merge to form the spinal nerves,
which consequently contain both sensory and motor fibres. As the spinal nerves
travel into the periphery they split into branches and the exact composition
of the nerve in terms of motor and sensory fibres is, of course, determined
by the structures the nerve will innervate.
One nerve fibre consists of an axon and its
nerve sheath. Each axon in the peripheral nervous
system is surrounded by a sheath of Schwann cells. An individual Schwann cell
may surround the axon for several hundred micrometers, and it may, in the
case of unmyelinated nerve fibers, surround up to 30 separate axons. The axons
are housed within infoldings of the Schwann cell cytoplasm and cell membrane,
the mesaxon  .
.
 In
the case of myelinated nerve fibres, Schwann cells form a sheath around one
axon and surround this axon with several double layers (up to hundreds) of
cell membrane. The myelin sheath formed by the Schwann cell insulates the
axon, improves its ability to conduct and, thus, provides the basis for the
fast saltatory transmission of impulses. Each
Schwann cell forms a myelin segment, in which the cell nucleus is located
approximately in the middle of the segment. The node
of Ranvier is the place along the course of the axon where two myelin
segments abut.
In
the case of myelinated nerve fibres, Schwann cells form a sheath around one
axon and surround this axon with several double layers (up to hundreds) of
cell membrane. The myelin sheath formed by the Schwann cell insulates the
axon, improves its ability to conduct and, thus, provides the basis for the
fast saltatory transmission of impulses. Each
Schwann cell forms a myelin segment, in which the cell nucleus is located
approximately in the middle of the segment. The node
of Ranvier is the place along the course of the axon where two myelin
segments abut.
Fibre types in peripheral nerves:
- Type A fibres (myelinated) are 4 - 20 µm
in diameter and conduct impulses at high velocities (15 - 120 m per second).
Examples: motor fibers, which innervate skeletal muscles, and sensory fibres.
- Type B fibres (myelinated) are 1 - 4 µm
in diameter and conduct impulses with a velocity of 3 - 14 m per second.
Example: preganglionic autonomic fibres.
- Type C fibres (unmyelinated) are 0.2 -
1 µm thick and conduct impulses at velocities ranging from 0.2 to
2 m per second. Examples: autonomic and sensory fibres.
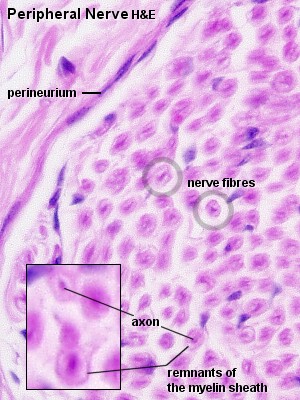
Peripheral nerves contain a considerable amount of connective tissue. The
entire nerve is surrounded by a thick layer of dense connective tissue, the
epineurium. Nerve fibres are frequently grouped
into distinct bundles, fascicles, within the
nerve. The layer of connective tissue surrounding the individual bundles is
called perineurium. The perineurium is formed
by several layers of flattened cells, which maintain the appropriate microenvironment
for the nerve fibres surrounded by them. The space between individual nerve
fibres is filled by loose connective tissue, the endoneurium.
Fibrocytes, macrophages and mast cells are present in the endoneurium.
Nerves are richly supplied by intraneural blood vessels, which form numerous
anastomoses. Arteries pass into the epineurium, form arteriolar networks in
the perineurium and give off capillaries to the endoneurium.
- Suitable Slides
- sections of peripheral
nerve - H&E, osmium
or plucked preparations of peripheral nerve - osmium
Peripheral Nerve, cat - osmium
Which structures can be recognized in peripheral nerves depends on the stain
that has been used in the preparation. Osmium gives a black color to lipids.
In osmium stained preparations it is possible to observe the myelin sheath
surrounding the axon. A good impression of the different sizes of the nerve
fibres may be obtained. The axon is usually not well preserved. It may only
form a little dark spot somewhere within the dark ring which represents the
myelin sheath. Lipid droplets in fat cells, which can be found in the connective
tissue around nerves, stand out as large (much larger than the nerve fibres),
round, homogeneously stained areas.
Draw the nerve at low magnification (you may include some
of the stained lipid droplets) and a small section of it at high magnification.


Peripheral Nerve, rat - H&E
In longitudinal H&E stained sections it is possible to identify
the axon running in its myelin sheath, nodes of Ranvier and Schwann cell nuclei.
Components of the connective tissue elements, which accompany the nerve, should
be visible and identifiable in both longitudinal and transverse sections.
H&E stained and transversely cut preparations give a good picture of the
axon in the middle of a ring-like structure (sometimes
fussy), which represents the remains of the myelin sheath. Due to their
small size and the lack of a myelin sheath, type C fibres are very difficult
to detect in either osmium or H&E stains.
Draw part of the longitudinally and transversely sectioned
nerve at high magnification. Include Schwann cell nuclei, myelin sheath, axons
and, if possible, nodes of Ranvier.

Ganglia are aggregations of nerve cells (ganglion cells) outside the CNS. Cranial nerve and dorsal root ganglia are surrounded by a connective tissue capsule, which is continuous with the dorsal root epi- and perineurium. Individual ganglion cells are surrounded by a layer of flattened satellite cells. Neurones in cranial nerve and dorsal root ganglia are pseudounipolar. They have a T-shaped process. The arms of the T represent branches of the neurite connecting the ganglion cell with the CNS (central branch) and the periphery (peripheral branch). Both branches function as one actively conducting axon, which
transmits information from the periphery to the CNS. The stem is connected to the perikaryon of the ganglion cell and is the only process originating from it. Ganglion cells in dorsal root ganglia do not receive synapses. Their function is the trophic support of their neurites.
Early in development two processes emerge from the perikaryon
of dorsal root ganglion cells, which merge in the course of development. These
ganglion cells are therefore also called pseudounipolar neurones. Two processes
emerge from the perikaryon of bipolar neurones. The majority of CNS neurones
are multipolar, i.e. more than two processes (but only one axon) emerge from
their perikaryon.
Autonomic ganglia do contain synapses, and the ganglion cells within them do have dendrites. They receive synapses from the first neurone of the two-neurone chain, which characterises most of the efferent connections of the autonomic nervous system. The second neurone is the ganglion cell itself. Some autonomic ganglia are embedded within the walls of the organs which they innervate (intramural ganglia - e.g. GIT and bladder).
- Suitable Slides
- sections of dorsal root
ganglia and autonomic ganglia - H&E,
toluidine blue, Giemsa, luxol fast blue/cresyl/violet (LFB/CV)
Dorsal Root Ganglion, cat - H&E and Autonomic
Ganglion - H&E
Ganglion cells will typically be several times larger than other cells
in the ganglia. The perikaryon is very large and surrounds a large and light
nucleus. Only the cells immediately surrounding the ganglion cells as one
flattened layer are satellite cells. With a lot of luck you may see the process
of a ganglion cell as it passes out of the capsule of satellite cells. Ganglion
cells are of course in contact with other parts of the nervous system and
with the peripheral tissues which they innervate. Consequently, nerve fibers
will be visible close to or within the ganglion.
Sketch the appearance of the spinal ganglion section at
low magnification. Draw a small section of the spinal ganglion and peripheral
ganglion at high magnification. Label ganglion cells, satellite cells and,
if visible, nerve fibres and connective tissue elements.


