Les tissus musculaires (deuxième partie)
5.4. Appareil de contraction
 L'appareil cellulaire de la contraction dépend du type
musculaire. Dans la fibre musculaire squelettique, il est scindé
en myofibrilles longitudinales et ce partage détermine
la striation longitudinale. Chaque myofibrille est une succession
de sarcomères. La disposition des myofibrilles dans le
cytoplasme est telle que les sarcomères sont rigoureusement
superposés, ce qui détermine la striation transversale.
Dans la fibre musculaire cardiaque, l'appareil contractile est
aussi composé de sarcomères mais ils ne sont pas
alignés en myofibrilles. La fibre musculaire lisse ne comprend
que des myofilaments épars dans le cytoplasme.
L'appareil cellulaire de la contraction dépend du type
musculaire. Dans la fibre musculaire squelettique, il est scindé
en myofibrilles longitudinales et ce partage détermine
la striation longitudinale. Chaque myofibrille est une succession
de sarcomères. La disposition des myofibrilles dans le
cytoplasme est telle que les sarcomères sont rigoureusement
superposés, ce qui détermine la striation transversale.
Dans la fibre musculaire cardiaque, l'appareil contractile est
aussi composé de sarcomères mais ils ne sont pas
alignés en myofibrilles. La fibre musculaire lisse ne comprend
que des myofilaments épars dans le cytoplasme.
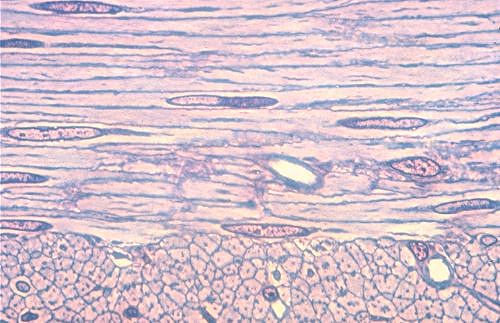
Dans cette fibre musculaire squelettique vue à fort grossissement,
l'appareil contractile occupe la quasi totalité du volume
cellulaire. Les stries longitudinales et transversales sont évidentes.
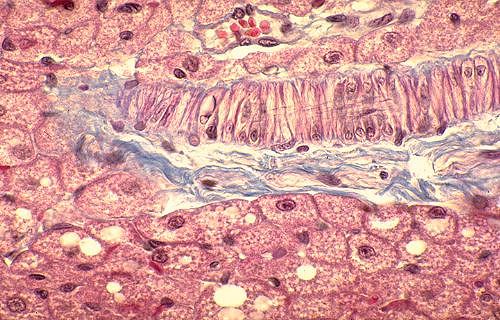
 Dans les fibres musculaires cardiaques, l'appareil contractile
est une masse fissurée. La fissure principale, centrale,
contient un noyau clair entouré d'organites cytoplasmiques.
La striation transversale est nette. Notez la présence
de capillaires dans les espaces de Henlé; les noyaux de
leur endothélium font saillie dans les lumières
vasculaires.
Dans les fibres musculaires cardiaques, l'appareil contractile
est une masse fissurée. La fissure principale, centrale,
contient un noyau clair entouré d'organites cytoplasmiques.
La striation transversale est nette. Notez la présence
de capillaires dans les espaces de Henlé; les noyaux de
leur endothélium font saillie dans les lumières
vasculaires.
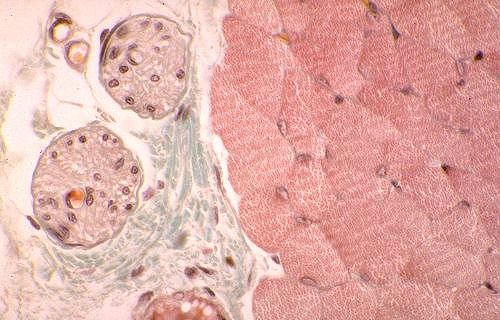
 Dans les fibres musculaires lisses, l'appareil contractile est
fait de myofilaments parallèles au grand axe de la cellule.
Ils sont bien visibles dans la coupe longitudinale, située
dans la partie supérieure de l'image.
Dans les fibres musculaires lisses, l'appareil contractile est
fait de myofilaments parallèles au grand axe de la cellule.
Ils sont bien visibles dans la coupe longitudinale, située
dans la partie supérieure de l'image.
Des corps denses ovoïdes ou fusiformes sont dispersés
entre les myofilaments et répartis à intervalles
réguliers sur la face interne de la membrane des cellules
musculaires lisses (flèches). Ils sont tous reliés
par des filaments intermédiaires, d'un diamètre
de 10 nm, et serviraient d'ancrage aux myofilaments d'actine.
Dans les trois types de fibres musculaires, les myofilaments
qui constituent l'appareil contractile comprennent des filaments
minces d'actine et des filaments épais de myosine.
Le diamètre des filaments d'actine, ou actine F, est de
5 nm et leur longueur de 1 µm. Ils sont composés de
deux chaînes tressées dont chaque spire s'étend
sur 40 nm. Chaque chaîne est une association de monomères
d'une protéine globuleuse, l'actine G.
Les filaments de myosine, d'un diamètre de 15 nm, sont
constitués de 300 à 400 molécules. Une molécule
de myosine, représentée en A, comprend deux chaînes
lourdes et quatre chaînes légères. Chaque
chaîne lourde est composée d'une tête globuleuse
et d'un segment hélicoïdal. A chaque tête sont
associées deux chaînes légères. Les
segments hélicoïdaux des deux chaînes lourdes
sont unis entre eux pour former une queue d'où émergent
les deux têtes. Dans un filament, représenté
en B, les molécules de myosine sont unies par leurs segments
hélicoïdaux et leurs têtes se projettent vers
l'extérieur au niveau de ses deux extrémités.
Dans les cellules musculaires lisses, les myofilaments sont dispersés.
Dans les cellules musculaires squelettiques et cardiaques, ils
sont organisés en sarcomères. Le sarcomère,
unité morphologique et fonctionnelle de l'appareil contractile,
s'étend entre deux lignes Z et mesure environ deux microns.
Dans cette micrographie électronique de cellules musculaires
squelettiques, les lignes Z sont indiquées par des flèches.
Elles sont très denses et partagent les bandes claires
I. Les bandes sombres A sont scindées par la bande H, elle-même
partagée par la ligne M. Chaque sarcomère est donc
constitué d'une demi-bande I, d'une bande A et d'une autre
demi-bande I.
Cette organisation en sarcomères provient de l'arrangement
des myofilaments épais et minces. La bande A est formée
de filaments épais de myosine. Ils sont unis par des ponts
transversaux au niveau de la ligne M, mais sont isolés
les uns des autres dans le reste de la bande A. Leur disposition
est très régulière et, en coupe transversale,
dessine un hexagone. En dehors de la bande H, les filaments de
myosine sont hérissés d'ergots ou têtes disposés
en spirale. Entre eux s'insinuent les filaments minces d'actine.
Six filaments d'actine sont disposés régulièrement
autour d'un filament de myosine. La bande I est formée
uniquement de filaments minces d'actine qui sont unis, entre sarcomères
voisins, au niveau de la ligne Z.
La contraction du muscle squelettique est le résultat du
glissement des filaments d'actine sur les filaments de myosine.
Ce glissement explique les changements de la myofibrille en fonction
de l'état de contraction ou d'étirement de la cellule
musculaire. Au cours de la contraction, les filaments d'actine
sont profondément enfoncés dans la bande A. La bande
I et la bande H sont alors très étroites. Au cours
de l'étirement, les filaments d'actine sont presque exclus
de la bande A. La bande I et la bande H sont alors très
larges. Au repos, les dimensions de la bande I et de la bande
H sont intermédiaires. Quel que soit l'état du muscle,
la longueur de la bande A est toujours la même.
La liaison de l'actine à la myosine, au cours de la contraction
et du relâchement, subit plusieurs modifications successives.
Dans une première étape, la tête de la myosine,
à laquelle sont liés un ADP et un phosphate, entre
en contact avec l'actine. Dans une deuxième étape,
l'ADP et le phosphate quittent la tête de la myosine qui
change d'orientation; dès lors, la molécule de myosine
tire le filament d'actine et le déplace dans le sens indiqué
par la flèche, vers le centre du sarcomère qui se
raccourcit. Dans une troisième étape, une molécule
d'ATP se lie à la tête de la myosine qui se détache
de l'actine. La quatrième étape clôture le
cycle : la molécule d'ATP est hydrolysée et la tête
de la myosine reprend son orientation initiale.
5.5. Jonctions neuromusculaires
Les jonctions neuromusculaires sont importantes car la plupart
des réflexes se traduisent finalement et quel que soit
leur point de départ par une réaction musculaire.
Les sensations recueillies au niveau des muscles déterminent
par ailleurs une série de réflexes posturaux et
autres, souvent caractéristiques de l'espèce.
Les fibres musculaires lisses et cardiaques sont innervées
par des terminaisons motrices nues, non myélinisées,
en provenance des plexus autonomiques. Dans le muscle squelettique,
il existe deux types de jonctions neuromusculaires, la plaque
motrice et les fuseaux neuromusculaires.
5.5.1. Plaque motrice
La plaque motrice est la jonction spécialisée entre
l'extrémité de l'axone d'un motoneurone alpha et
la fibre musculaire squelettique.
Avant d'atteindre la plaque motrice, l'axone perd sa gaine de
myéline et son endonèvre. Le périnèvre
s'arrime au sarcolemme. L'axone se ramifie en un grand nombre
de terminaisons dilatées. Chaque terminaison axonique dilatée
en bouton est riche en mitochondries et en vésicules contenant
un neurotransmetteur, l'acétylcholine. Elle est logée
dans une dépression de la cellule musculaire, mais reste
séparée du sarcolemme par une fente synaptique de
50 nm. La surface du sarcolemme est déprimée par
de nombreuses invaginations qui prolongent la fente synaptique
et contiennent une couche glycoprotéique. Au niveau de
la plaque motrice, la fibre musculaire contient de nombreuses
mitochondries et des noyaux groupés sous le sarcolemme.
La plaque motrice est effectrice. C'est une synapse chimique
où la terminaison axonique transmet l'influx en libérant
l'acétylcholine dans la fente synaptique et ses extensions.
L'acétylcholine se fixe aux récepteurs de la membrane
plasmique postsynaptique, ce qui accroît la perméabilité
du sarcolemme. L'entrée du sodium génère
un potentiel d'action qui se propage le long du sarcolemme.
L'acétylcholine libérée dans la fente synaptique
est détruite par l'acétylcholinestérase du
sarcolemme, ce qui bloque la réponse musculaire. La détection
de l'acétylcholinestérase par des méthodes
histochimiques permet de repérer les plaques motrices.
En voici une colorée en brun dans cette préparation.
5.5.2. Fuseau neuro-musculaire
 Les fuseaux neuromusculaires sont des organes récepteurs
situés dans la profondeur du muscle squelettique. Ils comprennent
des fibres musculaires modifiées appelées fibres
fusorielles, des terminaisons nerveuses sensitives encapsulées
avec support et des terminaisons nerveuses motrices.
Les fuseaux neuromusculaires sont des organes récepteurs
situés dans la profondeur du muscle squelettique. Ils comprennent
des fibres musculaires modifiées appelées fibres
fusorielles, des terminaisons nerveuses sensitives encapsulées
avec support et des terminaisons nerveuses motrices.
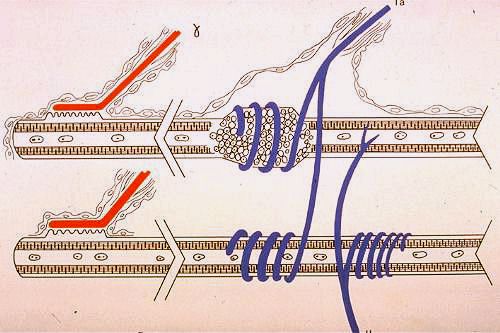
Voici le schéma d'un fuseau neuromusculaire. Les fibres
musculaires fusorielles sont différentes des autres fibres
musculaires. Certaines sont à chaîne nucléaire
et d'autres à poche nucléaire. Dans les fibres à
chaîne nucléaire, les noyaux sont centraux et les
myofibrilles sont périphériques. Dans les fibres
à poche nucléaire, la partie centrale de la fibre
est dilatée et cette dilatation est remplie d'une cinquantaine
de noyaux. Les fibres fusorielles sont en rapport étroit
avec des terminaisons nerveuses sensitives et motrices. Celles-ci,
en abordant le fuseau, perdent leur gaine de myéline et
leur endonèvre; leur périnèvre, qui forme
leur capsule, s'applique contre le sarcolemme de la fibre musculaire.
Les terminaisons sensitives prennent comme support la fibre fusorielle.
Celles de type Ia, ou primaires, s'enroulent en spirale autour
des poches nucléaires ou de la partie centrale des fibres
à chaîne nucléaire. Les terminaisons sensitives
de type II, ou secondaires, sont voisines des poches nucléaires
ou situées dans les parties juxta-médianes des fibres
à chaîne nucléaire. Deux terminaisons motrices
font partie du fuseau neuromusculaire. Des axones issus de motoneurones
alpha, et donc du même type que ceux destinés aux
cellules musculaires normales, se terminent à l'extrémité
des fibres fusorielles; ils ne sont pas représentés
dans le schéma. D'autres axones, provenant des motoneurones
gamma de la corne ventrale de la moelle épinière,
assurent l'innervation motrice propre au fuseau. Ils forment des
plaques motrices près des pôles des fibres fusorielles.
Voici un fuseau neuro-musculaire en coupe longitudinale. On y
reconnaît deux fibres fusorielles à poche nucléaire.
Des fibres nerveuses (flèches) entourées de leur
périnèvre aboutissent l'une au niveau de la poche
nucléaire, l'autre dans la partie polaire de la fibre fusorielle.
Les cellules périneurales, qui enveloppent le fuseau, sont
indiquées par des pointes de flèche.
Les fuseaux neuromusculaires font partie d'un ensemble de récepteurs
appelés propriocepteurs impliqués dans le contrôle
des mouvements et du tonus. Résumons leur rôle,
étudié plus en détail en physiologie. Les
fuseaux neuromusculaires interviennent dans le réflexe
myotatique qui contrôle de façon précise la
longueur des muscles tout particulièrement des muscles
étirés par la station debout. Les fibres fusorielles,
parallèles aux autres fibres musculaires, subissent le
même étirement que le reste du muscle. L'information
perçue par les terminaisons sensitives est transmise aux
motoneurones alpha dont les axones commandent la contraction musculaire.
L'innervation motrice propre au fuseau contrôle le niveau
de sensibilité du réflexe myotatique. Son excitation,
en contractant les extrémités polaires du fuseau
neuromusculaire, étire la zone équatoriale réceptrice
et augmente ainsi sa sensibilité.
5.6. Tissu nodal du myocarde
Le tissu nodal est l'ensemble des fibres musculaires cardiaques
modifiées qui stimulent la contraction cardiaque et conduisent
l'influx aux différentes parties du myocarde.
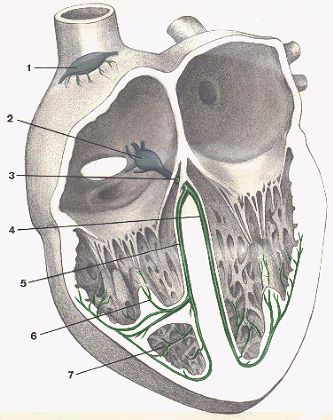
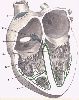 Anatomiquement, le tissu nodal est réparti en noeuds,
en faisceaux et en fibres. Le noeud le plus important est
le noeud sino-auriculaire de Keith et Flack (1), situé
près de l'orifice de la veine cave supérieure. C'est
dans ce noeud que naît le potentiel d'action. Il est transmis
au noeud auriculo-ventriculaire de Tawara (2) situé près
de l'orifice du sinus coronaire. Du noeud de Tawara part le faisceau
de His (3) qui transmet le potentiel d'action des oreillettes
vers les ventricules et est situé dans la partie supérieure
de la cloison interventriculaire. Ce faisceau se divise en deux
branches (4 et 5) destinées chacune à un ventricule.
Ces branches se ramifient en dessous de l'endocarde qui tapisse
les cavités cardiaques et se distribuent aux muscles papillaires
(6) et au myocarde. Ce réseau sous-endocardique est connu
sous le nom de cellules de Purkinje (7).
Anatomiquement, le tissu nodal est réparti en noeuds,
en faisceaux et en fibres. Le noeud le plus important est
le noeud sino-auriculaire de Keith et Flack (1), situé
près de l'orifice de la veine cave supérieure. C'est
dans ce noeud que naît le potentiel d'action. Il est transmis
au noeud auriculo-ventriculaire de Tawara (2) situé près
de l'orifice du sinus coronaire. Du noeud de Tawara part le faisceau
de His (3) qui transmet le potentiel d'action des oreillettes
vers les ventricules et est situé dans la partie supérieure
de la cloison interventriculaire. Ce faisceau se divise en deux
branches (4 et 5) destinées chacune à un ventricule.
Ces branches se ramifient en dessous de l'endocarde qui tapisse
les cavités cardiaques et se distribuent aux muscles papillaires
(6) et au myocarde. Ce réseau sous-endocardique est connu
sous le nom de cellules de Purkinje (7).
Les fibres musculaires du tissu nodal sont globuleuses. Leur noyau
central est entouré d'une zone claire. Les myofibrilles
sont disposées en périphérie contre la membrane
plasmique.
Vous terminerez votre étude en analysant les quatre dernières
images. Quels tissus pouvez-vous y identifier et pourquoi?
 Réponse
Réponse
 Réponse
Réponse
 Réponse
Réponse
 Réponse
Réponse
 L'appareil cellulaire de la contraction dépend du type
musculaire. Dans la fibre musculaire squelettique, il est scindé
en myofibrilles longitudinales et ce partage détermine
la striation longitudinale. Chaque myofibrille est une succession
de sarcomères. La disposition des myofibrilles dans le
cytoplasme est telle que les sarcomères sont rigoureusement
superposés, ce qui détermine la striation transversale.
Dans la fibre musculaire cardiaque, l'appareil contractile est
aussi composé de sarcomères mais ils ne sont pas
alignés en myofibrilles. La fibre musculaire lisse ne comprend
que des myofilaments épars dans le cytoplasme.
L'appareil cellulaire de la contraction dépend du type
musculaire. Dans la fibre musculaire squelettique, il est scindé
en myofibrilles longitudinales et ce partage détermine
la striation longitudinale. Chaque myofibrille est une succession
de sarcomères. La disposition des myofibrilles dans le
cytoplasme est telle que les sarcomères sont rigoureusement
superposés, ce qui détermine la striation transversale.
Dans la fibre musculaire cardiaque, l'appareil contractile est
aussi composé de sarcomères mais ils ne sont pas
alignés en myofibrilles. La fibre musculaire lisse ne comprend
que des myofilaments épars dans le cytoplasme.