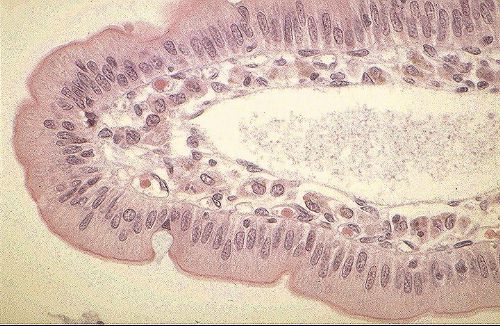
Le système circulatoire (cours 2)
 Les capillaires relient les artérioles aux veinules,
faisant ainsi de l'appareil vasculaire sanguin un système
continu. Tandis que les artères et les veines appartiennent
au réseau de la macrocirculation et constituent des voies
de transit, les capillaires font partie de la microcirculation
et sont le siège d'échanges métaboliques
entre le sang et les tissus voisins.
Les capillaires relient les artérioles aux veinules,
faisant ainsi de l'appareil vasculaire sanguin un système
continu. Tandis que les artères et les veines appartiennent
au réseau de la macrocirculation et constituent des voies
de transit, les capillaires font partie de la microcirculation
et sont le siège d'échanges métaboliques
entre le sang et les tissus voisins.
Le réseau microcirculatoire
ou lit vasculaire périphérique comprend plusieurs
éléments. Il commence par l'artériole
(1) que nous avons décrite. La métartériole
(2) dérive de l'artériole et en a la structure.
Elle se continue par une veinule (3) dont le diamètre
est double ou triple. Sur la métartériole se branchent
des capillaires (4) et, au niveau de leur jonction, se
trouvent les sphincters précapillaires (5).
A la jonction des capillaires avec la métartériole,
la paroi vasculaire ressemble à celle d'une artériole.
Elle comprend, en effet, une couche de cellules endothéliales
(1) et une couche de cellules musculaires lisses (2) séparées
par la membrane basale (3). Parfois, quelques évaginations
du cytoplasme endothélial (4) refoulent la face interne
des cellules musculaires lisses. Les cellules musculaires lisses
du sphincter sont innervées par des nerfs amyélinisés
et les terminaisons renflées des axones se logent
dans une invagination de leur cytoplasme (5).
En se relâchant ou en se contractant, les sphincters précapillaires
règlent le flux sanguin dans les capillaires.
Cette fonction est contrôlée par le système
nerveux autonomique mais est aussi influencée par des médiateurs
chimiques. L'adrénaline provoque la constriction des sphincters
et empêche donc l'arrivée du sang dans les capillaires;
l'histamine, l'acétylcholine, les kinines vasoactives provoquent
le relâchement des sphincters et donc la dilatation des
capillaires.
L'activité des sphincters est également influencée
par le métabolisme local puisqu'une diminution de la pression
partielle en oxygène dans une région y provoque
leur relâchement et donc un afflux sanguin.
Les capillaires sont des segments
vasculaires très grêles dont le diamètre est
compris entre 3 et 10 micromètres. Leur calibre correspond
donc approximativement au diamètre d'un globule rouge.
Leur paroi est formée exclusivement d'un endothélium
et de sa membrane basale.
On distingue parmi eux : les capillaires continus, les capillaires
fenestrés et les sinusoïdes.
 Dans les capillaires continus l'endothélium qui
limite la lumière centrale ne présente aucune discontinuité.
Les cellules endothéliales sont aplaties et leurs noyaux
font saillie dans la lumière.
Dans les capillaires continus l'endothélium qui
limite la lumière centrale ne présente aucune discontinuité.
Les cellules endothéliales sont aplaties et leurs noyaux
font saillie dans la lumière.
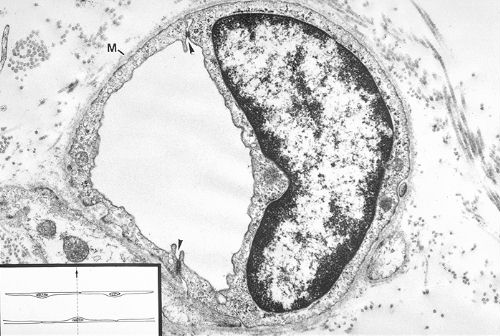
Voici ce capillaire continu dans une micrographie électronique.
La coupe traverse deux cellules mais un seul noyau où le
nucléole est peu apparent et l'hétérochromatine
disposée le long de la membrane nucléaire.
Le cytoplasme contient certes les organites habituels mais surtout
de nombreuses petites vésicules.
Les cellules endothéliales sont unies par des jonctions
indiquées ici par des flèches. Leur face externe
est revêtue d'une membrane basale continue (M). Des jonctions
communicantes servent aux échanges entre cellules.
Voici à plus fort grossissement le cytoplasme d'une autre cellule endothéliale.
Les vésicules y sont très nombreuses. Leur diamètre
est variable, mais mesure habituellement 60 à 70 nanomètres.
Ces vésicules se forment à partir d'invaginations
de la membrane plasmique et s'ouvrent des deux côtés
de la cellule endothéliale. Il s'agit de vésicules
de diacytose servant au passage transcellulaire de substances.
La flèche désigne un couloir formé par la
fusion de plusieurs vésicules.
L'endothélium des capillaires est souvent entouré
de quelques cellules isolées appelées péricytes
et situées en dehors de la membrane basale. Ce sont des
cellules allongées possédant un large noyau. Leur
cytoplasme est riche en mitochondries et surtout en réticulum
endoplasmique rugueux.
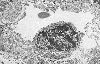 Les capillaires fenestrés
se trouvent notamment dans les glomérules rénaux
et les glandes endocrines. Leur endothélium est
très mince et perforé de pores d'environ 100 nanomètres.
Les capillaires fenestrés
se trouvent notamment dans les glomérules rénaux
et les glandes endocrines. Leur endothélium est
très mince et perforé de pores d'environ 100 nanomètres.
Dans les capillaires des glomérules rénaux, les
pores sont ouverts; mais dans les capillaires des glandes endocrines,
ils sont fermés par une fine membrane unitaire ou diaphragme
désigné ici par des flèches.
Les pores sont bien évidents lorsque la membrane plasmique
est étudiée en cryofracture; voici une portion de cette membrane plasmique limitant un capillaire fenestré.
Dans la lumière, le globule rouge est marqué d'un
astérisque. Les pores sont circulaires et uniformément
répartis. Leur taille est régulière.
Les capillaires sont le siège des échanges métaboliques
entre le sang et les tissus voisins. Plusieurs voies d'échanges
ont été proposées.
L'eau, les substances de faible poids moléculaire ou liposolubles
traversent directement l'endothélium par diffusion.
D'autres molécules plus volumineuses sont endocytées
au niveau d'une face cellulaire, traversent la cellule par diacytose
et sont exocytées au niveau de la face opposée;
parfois les vésicules d'endocytose fusionnent et forment
un véritable couloir d'une face cellulaire à l'autre.
Les mêmes modes d'échange existent au niveau des
capillaires fenestrés mais les pores y constituent
une voie supplémentaire de transit. Le transport via les
fenestrations fermées dépend de la perméabilité
du diaphragme, qui n'empêche pas le passage de molécules
telles que la ferritine ou le dextran. La perméabilité
de la membrane basale formée de mucopolysaccharides
et de glycoprotéines pourrait également régler
la vitesse des échanges.
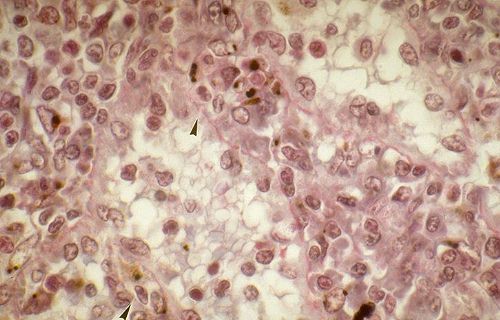
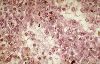 Les sinusoïdes sanguins
sont très larges. Leur paroi est percée d'orifices
parce qu'il existe des interstices entre les cellules endothéliales.
Ils sont dépourvus de membrane basale. Voici des sinusoïdes
dans la rate. On les reconnaît à l'importance de
leur lumière très irrégulière, limitée
par les cellules endothéliales. Les interstices entre les
cellules endothéliales sont indiqués par des flèches.
Les sinusoïdes sanguins
sont très larges. Leur paroi est percée d'orifices
parce qu'il existe des interstices entre les cellules endothéliales.
Ils sont dépourvus de membrane basale. Voici des sinusoïdes
dans la rate. On les reconnaît à l'importance de
leur lumière très irrégulière, limitée
par les cellules endothéliales. Les interstices entre les
cellules endothéliales sont indiqués par des flèches.
La discontinuité de l'endothélium est bien visible
lorsque l'on examine la face externe d'un sinusoïde au microscope à balayage.
La paroi de l'endothélium est percée d'orifices
au point d'être réduite à un grillage. A travers
les orifices, on aperçoit la cavité du sinusoïde
et les globules rouges qu'il contient.
 L'union de plusieurs capillaires donne des veinules qui
ellesmêmes se déversent dans des veines de
diamètre croissant. Habituellement on classe les veines
en fonction de leur taille. Dans l'espace humaine, les petites
veines ont un diamètre compris entre 50 micromètres
et 1 mm; les veines moyennes ont un diamètre compris
entre 1 et 10 mm et les larges veines ont plus d'un cm
de diamètre.
L'union de plusieurs capillaires donne des veinules qui
ellesmêmes se déversent dans des veines de
diamètre croissant. Habituellement on classe les veines
en fonction de leur taille. Dans l'espace humaine, les petites
veines ont un diamètre compris entre 50 micromètres
et 1 mm; les veines moyennes ont un diamètre compris
entre 1 et 10 mm et les larges veines ont plus d'un cm
de diamètre.
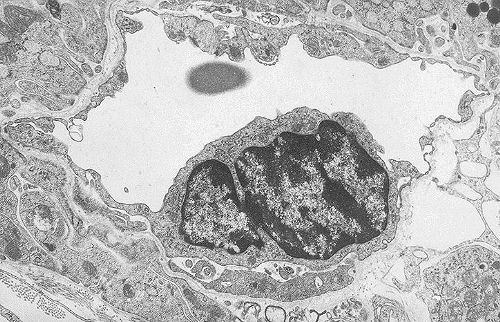
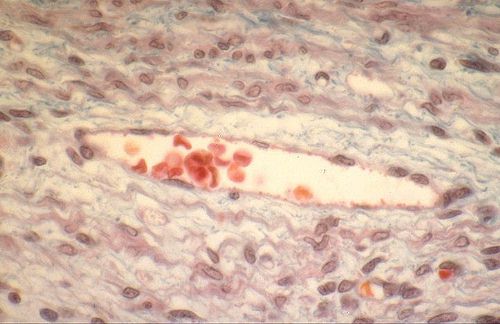
Les veinules ont le même aspect que les capillaires, mais
leur diamètre est supérieur. Voici une veinule dans une coupe de tissu conjonctif.
Sa paroi est très mince et formée exclusivement
d'un endothélium.
Comme les capillaires, les veinules interviennent dans les échanges
métaboliques. Leur principale caractéristique est
leur sensibilité aux agents inflammatoires qui augmentent
leur perméabilité, ce qui permet la diffusion du
plasma et le passage de protéines de haut poids moléculaire.
Ces agents favorisent aussi l'adhérence des leucocytes
aux parois vasculaires et leur diapédèse, en ralentissant
le flux sanguin.
De nombreuses veines petites et moyennes, et principalement celles
des extrémités sont munies de valvules
qui empêchent le reflux.
L'axe conjonctif de ces valvules se projette dans la lumière
toujours dans la direction du sang. Il est recouvert par l'endothélium.
La paroi des veines larges et moyennes
est mince comparée à celle des artères correspondantes
et elle ne peut être subdivisée nettement en intima,
média et adventice.
Voici une veine fémorale de souris.
L'endothélium limite la cavité. Le reste de la paroi
est formé de tissu conjonctif, où sont dispersées
des cellules musculaires lisses concentriques à la lumière.
Elles ne forment pas de couches régulières et continues
comme dans les artères.
 Généralement, un réseau de capillaires connecte
les artères et les veines. Ce schéma est différent
dans certains organes. Ainsi dans le foie et l'hypophyse,
la circulation est du type porte : un réseau de
capillaires (1 et 3) est situé entre deux veines (2 et
4).
Généralement, un réseau de capillaires connecte
les artères et les veines. Ce schéma est différent
dans certains organes. Ainsi dans le foie et l'hypophyse,
la circulation est du type porte : un réseau de
capillaires (1 et 3) est situé entre deux veines (2 et
4).
Dans les glomérules rénaux,
la circulation est du type admirable.
Une artériole afférente (1) se divise en
capillaires (2); ceuxci se regroupent pour former l'artériole
efférente (3).
La disposition classique artèrescapillairesveines
peut également être modifiée par l'existence
de communications directes entre les artères et
les veines. Cellesci peuvent être simples ou former
de petits organes complexes, les glomi vasculaires
que l'on trouve notamment dans le derme.
Dans les glomi, l'anastomose artérioveineuse est
pelotonnée sur ellemême et plusieurs segments
de l'artériole et de la veine sont donc visibles dans une
seule coupe. La paroi de la partie artériolaire est très
épaisse et formée de cellules épithélioïdes.
Ce sont des cellules musculaires modifiées dont la disposition
rappelle celle d'un épithélium mais qui gardent
la capacité de se contracter. La partie veineuse est large
et sa paroi est très mince.
Les glomi sont délimités par une capsule de tissu
conjonctif dense. Ils sont richement innervés par le système
nerveux sympathique. Ils règlent le flux sanguin et
interviennent dans la thermorégulation. Les corpuscules
carotidiens et aortiques ont une structure apparentée à
celle des glomi.
 Le système lymphatique
draine la lymphe vers la circulation sanguine. La lymphe est
le liquide interstitiel continuellement renouvelé par les
capillaires; elle est formée d'eau, d'électrolytes,
de protéines et, pour la circulation intestinale, de lipides.
Elle contient de nombreux lymphocytes. Le liquide interstitiel
est drainé par les capillaires lymphatiques qui déversent
leur contenu dans des vaisseaux lymphatiques plus importants.
Ceuxci aboutissent aux ganglions lymphatiques où
la lymphe est filtrée. De là, elle passe dans le
canal thoracique ou le canal lymphatique droit qui se jettent
dans le confluent jugulosousclavier.
Le système lymphatique
draine la lymphe vers la circulation sanguine. La lymphe est
le liquide interstitiel continuellement renouvelé par les
capillaires; elle est formée d'eau, d'électrolytes,
de protéines et, pour la circulation intestinale, de lipides.
Elle contient de nombreux lymphocytes. Le liquide interstitiel
est drainé par les capillaires lymphatiques qui déversent
leur contenu dans des vaisseaux lymphatiques plus importants.
Ceuxci aboutissent aux ganglions lymphatiques où
la lymphe est filtrée. De là, elle passe dans le
canal thoracique ou le canal lymphatique droit qui se jettent
dans le confluent jugulosousclavier.
Les capillaires lymphatiques se trouvent partout sauf dans le
système nerveux central, les épithéliums,
le cartilage, la moelle osseuse et le thymus. Ils débutent
en culdesac, sont de calibre très variable
et forment un réseau irrégulier. Leur paroi comprend
une couche unique et continue de cellules endothéliales.
La lumière ne contient jamais de globules rouges mais
parfois un précipité amorphe de protéines
qui se forme lorsque la préparation est fixée et
déshydratée.
Les cellules endothéliales des capillaires lymphatiques
sont très minces. Leur noyau est allongé. Leur cytoplasme
contient tous les organites habituels mais est surtout caractéristique
par sa richesse en vésicules de micropinocytose.
Il n'existe ni jonctions spécialisées entre cellules
ni membrane basale.
La cellule endothéliale représentée dans cette micrographie
est séparée de la cellule glandulaire sousjacente
par quelques fibres de collagène.
Les vaisseaux lymphatiques recueillent la lymphe drainée
par les capillaires lymphatiques. Ils sont souvent munis de valvules.
Cellesci sont formées de deux couches endothéliales
séparées par une fine bande de tissu conjonctif.
Elles se projettent dans la lumière du vaisseau et sont
toujours orientées dans la direction du flux lymphatique.
 Les capillaires relient les artérioles aux veinules,
faisant ainsi de l'appareil vasculaire sanguin un système
continu. Tandis que les artères et les veines appartiennent
au réseau de la macrocirculation et constituent des voies
de transit, les capillaires font partie de la microcirculation
et sont le siège d'échanges métaboliques
entre le sang et les tissus voisins.
Les capillaires relient les artérioles aux veinules,
faisant ainsi de l'appareil vasculaire sanguin un système
continu. Tandis que les artères et les veines appartiennent
au réseau de la macrocirculation et constituent des voies
de transit, les capillaires font partie de la microcirculation
et sont le siège d'échanges métaboliques
entre le sang et les tissus voisins.